
Lab 14: Urinary system & Reproductive system
When you are prepared for the section of the Lab Practical on Week 14 Learning Objectives in Week 15, you will be able to:
- Identify organs of the urinary system and their functions.
- Identify regions of the kidney and of the nephron and how they are involved in urine production.
- Identify parts of the male reproductive system and describe their functions.
- Identify parts of the female reproductive system and describe their functions.
- Identify structures within the ovary and describe how they are connected to oocyte maturation.
- Describe the pathway of gametes as they travel through male and female bodies.
The Urinary system
You are likely familiar with the urinary system’s role in cleansing the blood and ridding the body of wastes through the production of urine, but there are many other important functions that the urinary system participates in. The urinary system plays a role in regulation of pH, blood pressure, the concentration of solutes in the blood, and the concentration of red blood cells. The kidneys also perform the final synthesis step of vitamin D production, converting calcidiol to calcitriol, the active form of vitamin D.
Urine is a fluid of variable composition that requires specialized structures to remove it from the body safely and efficiently. Blood is filtered, and the filtrate is transformed into urine at a relatively constant rate throughout the day in the kidneys. This processed liquid is transported via the ureters to the urinary bladder, where it is stored until a convenient time for excretion. Urine then exits the body via the urethra. All structures involved in the transport and storage of the urine are large enough to be visible to the naked eye (Figure 14.1).
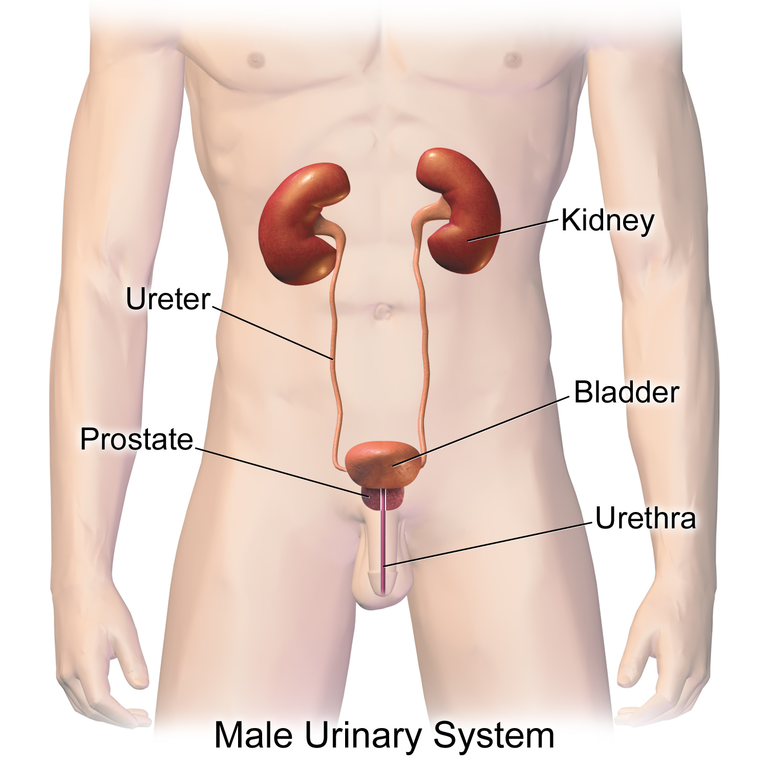
This transport and storage system not only stores the waste, but it protects the tissues from damage due to the wide range of pH and osmolarity of the urine, prevents infection by foreign organisms, and for the male, provides reproductive functions.
Kidneys
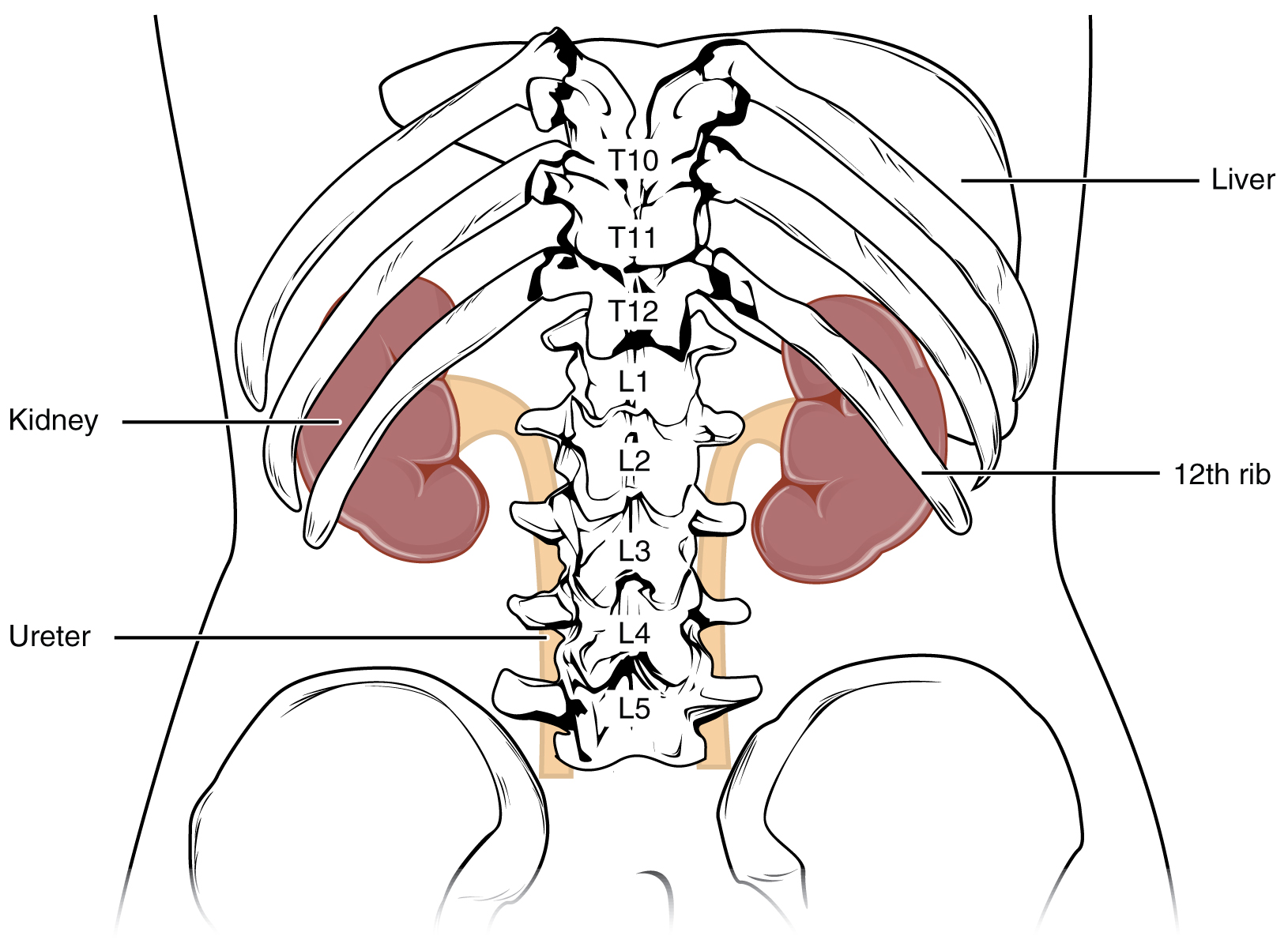
The kidneys lie on either side of the spine in the retroperitoneal space between the parietal peritoneum and the posterior abdominal wall, well protected by muscle, fat, and ribs. The left kidney is located at about the T12 to L3 vertebrae, whereas the right is lower due to slight displacement by the liver (Figure 14.2). They are roughly the size of your fist. The male kidney is typically a bit larger than the female kidney. The kidneys are well vascularized, receiving about twenty-five percent of the cardiac output at rest.
A frontal section through the kidney reveals an outer region called the renal cortex and an inner region called the medulla (Figure 14.3). The renal columns are connective tissue extensions that radiate downward from the cortex through the medulla to separate the most characteristic features of the medulla, the renal pyramids (medullary pyramids) and renal papillae. The papillae are bundles of collecting ducts that transport urine made by nephrons to the calyces of the kidney for excretion. The renal columns also serve to divide the kidney into 6–8 lobes and provide a supportive framework for vessels that enter and exit the cortex. The pyramids and renal columns taken together constitute the kidney lobes.
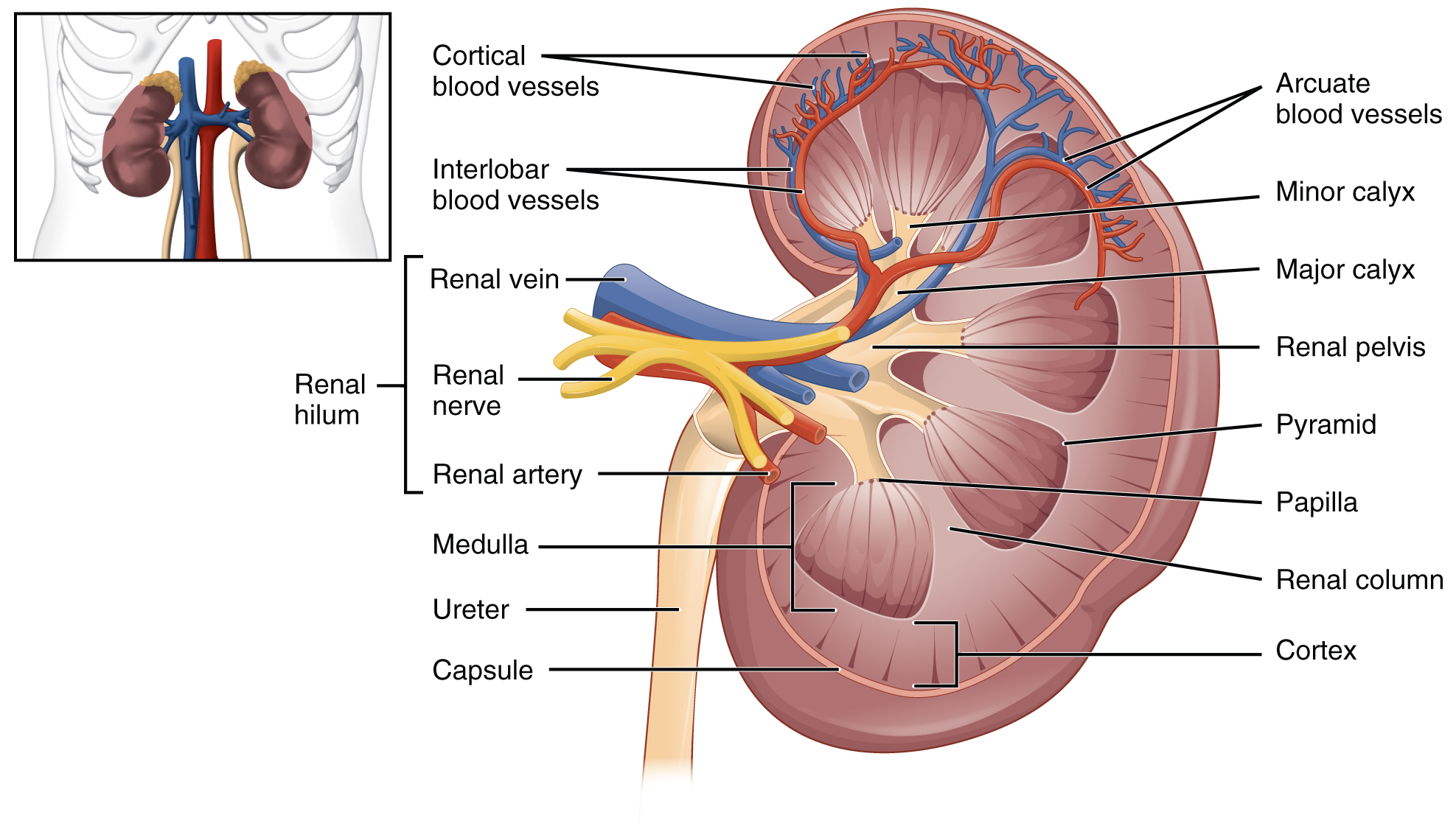
The renal hilum is the entry and exit site for structures servicing the kidneys: vessels, nerves, lymphatics, and ureters. The medial-facing hila are tucked into the sweeping convex outline of the cortex. Emerging from the hilum is the renal pelvis, which is formed from the major and minor calyxes in the kidney. The smooth muscle in the renal pelvis funnels urine via peristalsis into the ureter. The renal arteries form directly from the abdominal aorta, whereas the renal veins return cleansed blood directly to the inferior vena cava. The artery, vein, and renal pelvis are arranged in an anterior-to-posterior order.
Nephrons and Vessels
The renal artery first divides into segmental arteries, followed by further branching to form interlobar arteries that pass through the renal columns to reach the cortex (Figure 14.4). Smaller arteries branch from the interlobar arteries until eventually the blood reaches the afferent arterioles. The afferent arterioles service about 1.3 million nephrons in each kidney.
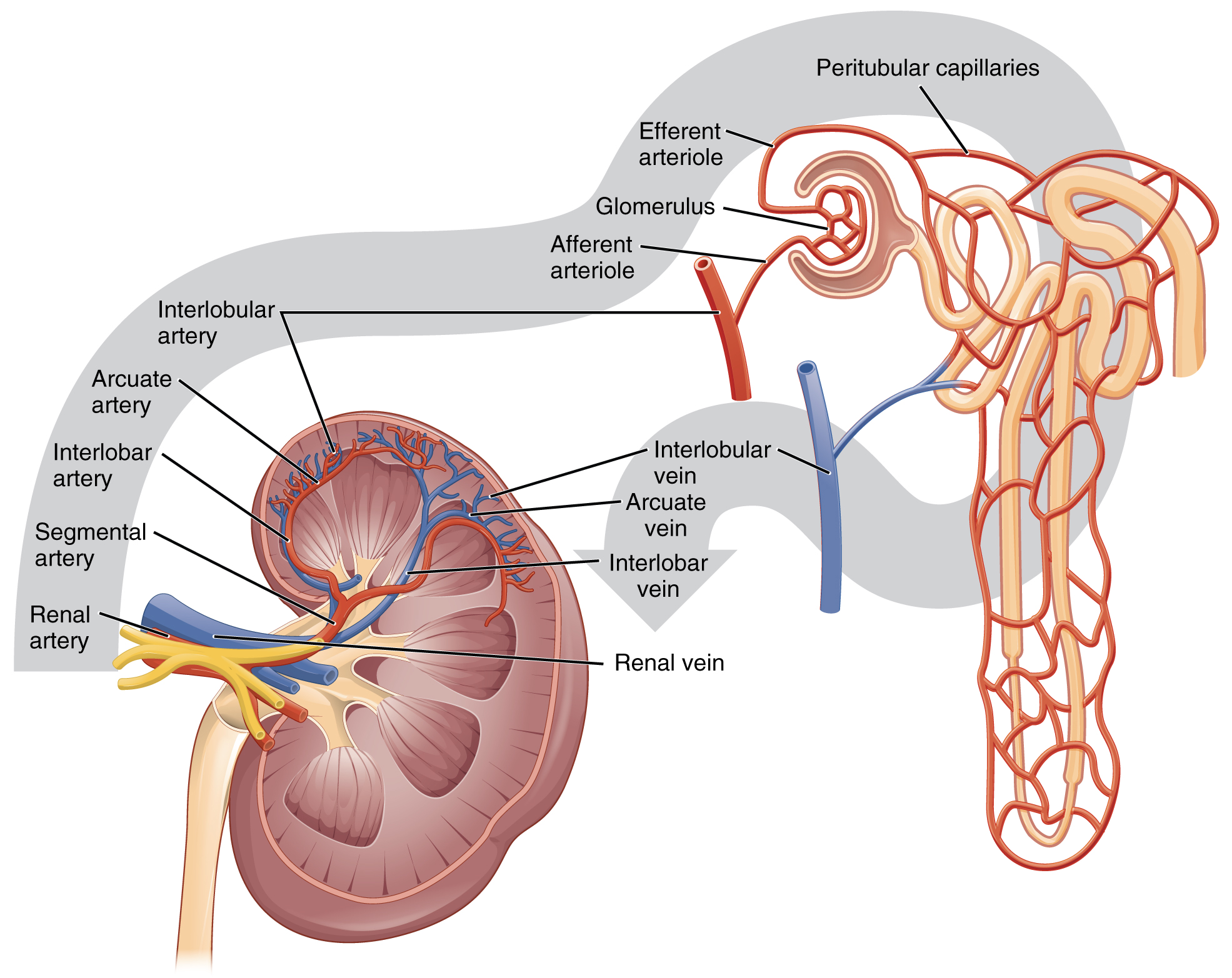
Nephrons are the “functional units” of the kidney; they cleanse the blood and balance the constituents of the circulation. The afferent arterioles form a tuft of high-pressure capillaries about 200 µm in diameter, the glomerulus. The rest of the nephron consists of a continuous tubule whose proximal end surrounds the glomerulus as a glomerular capsule (Bowman’s capsule). The glomerulus and glomerular capsule together form the renal corpuscle. After passing through the renal corpuscle, the capillaries form a second arteriole, the efferent arteriole (Figure 14.5). These will next form a capillary network around the more distal portions of the nephron tubule, the peritubular capillaries and vasa recta, before returning to the venous system. As the glomerular filtrate progresses through the nephron, these capillary networks recover most of the solutes and water, and return them to the circulation. Since a capillary bed (the glomerulus) drains into a vessel that in turn forms a second capillary bed, the definition of a portal system is met. This is the only portal system in which an arteriole is found between the first and second capillary beds. (Portal systems also link the hypothalamus to the anterior pituitary, and the blood vessels of the digestive viscera to the liver).
Fluid that leaks out of the glomerulus and is captured by the glomerular capsule is called filtrate. As the filtrate passes through the nephron, it is modified into urine. Nephrons accomplish this with three principle functions—filtration, reabsorption, and secretion. Many changes take place in the different parts of the nephron before urine is created for disposal. Filtered fluid collected by the glomerular capsule first travels to the proximal convoluted tubule. It is called convoluted due to its tortuous path. The fluid then goes to the descending and ascending portions of the loop of Henle (sometimes referred to as the nephron loop), which then leads to the final part of the nephron, the distal convoluted tubule (Figure 14.5).
Much of the nephron structures are located in the cortex of the kidney. In a dissected kidney, it is easy to identify the cortex; it appears lighter in color compared to the rest of the kidney. All of the renal corpuscles as well as both the proximal convoluted tubules (PCTs) and distal convoluted tubules are found here. Some nephrons have a short loop of Henle that does not dip beyond the cortex. These nephrons are called cortical nephrons. About 15 percent of nephrons have long loops of Henle that extend deep into the medulla and are called juxtamedullary nephrons.
The collecting ducts are continuous with the nephron but not technically part of it. In fact, each duct collects filtrate from several nephrons for final modification. Collecting ducts merge as they descend deeper in the medulla to form about 30 terminal ducts, which empty at a papilla. Once the fluid passes through the renal papilla, it is no longer modified and it is classified as urine.
Ureters
As urine is formed, it drains into the calyces of the kidney, which merge to form the funnel-shaped renal pelvis in the hilum of each kidney. The hilum narrows to become the ureter of each kidney. As urine passes through the ureter, it does not passively drain into the bladder but rather is propelled by waves of peristalsis. The ureters are approximately 30 cm long. The inner mucosa is lined with transitional epithelium and scattered goblet cells that secrete protective mucus. The muscular layer of the ureter consists of longitudinal and circular smooth muscles that create the peristaltic contractions to move the urine into the bladder without the aid of gravity (Figure 14.6). Finally, a loose adventitial layer composed of collagen and fat anchors the ureters between the parietal peritoneum and the posterior abdominal wall. The kidneys and ureters are completely retroperitoneal, and the bladder has a peritoneal covering only over the dome.
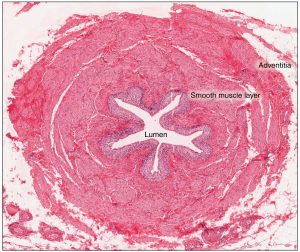
Urinary Bladder
The urinary bladder collects urine from both ureters (Figure 14.7). The bladder lies anterior to the uterus in females, posterior to the pubic bone and anterior to the rectum. During late pregnancy, its capacity is reduced due to compression by the enlarging uterus, resulting in increased frequency of urination. In males, the anatomy is similar, minus the uterus, and with the addition of the prostate inferior to the bladder. The bladder is partially retroperitoneal (outside the peritoneal cavity) with its peritoneal-covered “dome” projecting into the abdomen when the bladder is distended with urine.
The bladder is a highly distensible organ comprised of irregular crisscrossing bands of smooth muscle collectively called the detrusor muscle. The interior surface is made of transitional cellular epithelium that is structurally suited for the large volume fluctuations of the bladder. When empty, it resembles columnar epithelia, but when stretched, it “transitions” (hence the name) to a squamous appearance. Volumes in adults can range from nearly zero to 500–600 mL.
Urethra
The urethra transports urine from the bladder to the outside of the body for disposal. The urethra is the only urologic organ that shows any significant anatomic difference between males and females; all other urine transport structures are identical (Figure 14.8).
The urethra in both males and females begins inferior and central to the two ureteral openings forming the three points of a triangular-shaped area at the base of the bladder called the trigone (Greek tri- = “triangle”). The urethra tracks posterior and inferior to the pubic symphysis. In both males and females, the proximal urethra is lined by transitional epithelium, whereas the terminal portion is a nonkeratinized, stratified squamous epithelium. In the male, pseudostratified columnar epithelium lines the urethra between these two cell types. Voiding is regulated by an involuntary autonomic] nervous system-controlled internal urinary sphincter, consisting of smooth muscle and voluntary skeletal muscle that forms the external urinary sphincter below it.
Female Urethra
The female external urethral orifice is embedded in the anterior vaginal wall inferior to the clitoris, superior to the vaginal opening (introitus), and medial to the labia minora. The short length of the female urethra, about 4 cm, is less of a barrier to fecal bacteria than the longer male urethra and the best explanation for the greater incidence of UTI in women.
Male Urethra
The male urethra passes through the prostate gland immediately inferior to the bladder before passing below the pubic symphysis. The length of the male urethra varies between men but averages 20 cm in length. It is divided into three regions: the prostatic urethra, the membranous urethra, and the spongy (penile) urethra. The prostatic urethra passes through the prostate gland. During sexual intercourse, it receives sperm via the ejaculatory ducts and secretions from the seminal vesicles. The membranous urethra passes through the deep muscles of the perineum, where it is invested by the overlying urethral sphincters. The spongy urethra exits at the tip (external urethral orifice) of the penis after passing through the corpus spongiosum.
The Reproductive System
Small, uncoordinated, and slick with amniotic fluid, a newborn encounters the world outside of her mother’s womb. We do not often consider that a child’s birth is proof of the healthy functioning of both her mother’s and father’s reproductive systems. Moreover, her parents’ endocrine systems had to secrete the appropriate regulating hormones to induce the production and release of unique male and female gametes, reproductive cells containing the parents’ genetic material (one set of 23 chromosomes). Her parent’s reproductive behavior had to facilitate the transfer of male gametes—the sperm—to the female reproductive tract at just the right time to encounter the female gamete, an oocyte (egg). Finally, combination of the gametes (fertilization) had to occur, followed by implantation and development.
The Male Reproductive System
Unique for its role in human reproduction, a gamete is a specialized sex cell carrying 23 chromosomes—one half the number in body cells. At fertilization, the chromosomes in one male gamete, called a sperm (or spermatozoon), combine with the chromosomes in one female gamete, called an oocyte. The function of the male reproductive system is to produce sperm and transfer them to the female reproductive tract. The paired testes are a crucial component in this process, as they produce both sperm and androgens, the hormones that support male reproductive physiology. In humans, the most important male androgen is testosterone. Several accessory organs and ducts aid the process of sperm maturation and transport the sperm and other seminal components to the penis, which delivers sperm to the female reproductive tract.

Scrotum
The testes are located in a skin-covered, highly pigmented, muscular sack called the scrotum that extends from the body behind the penis. This location is important in sperm production, which occurs within the testes, and proceeds more efficiently when the testes are kept 2 to 4°C below core body temperature.
The dartos muscle and cremaster muscles, part of the walls of the scrotum, can contract to elevate the testes in cold weather (or water), moving the testes closer to the body and decreasing the surface area of the scrotum to retain heat. Alternatively, as the environmental temperature increases, the scrotum relaxes, moving the testes farther from the body core and increasing scrotal surface area, which promotes heat loss. Externally, the scrotum has a raised medial thickening on the surface called the raphae.

Testes
The testes (singular = testis) are the male gonads—that is, the male reproductive organs. They produce both sperm and androgens, such as testosterone, and are active throughout the reproductive lifespan of the male.
Paired ovals, the testes are each approximately 4 to 5 cm in length and are housed within the scrotum (Figure 14.10). They are surrounded by two distinct layers of protective connective tissue, the tunica vaginalis and the tunica albuginea. The tunica albuginea is a tough, white, dense connective tissue layer that covers the the outside of the testis, as well as invaginating to form septa that divide the testis into 300 to 400 structures called lobules. Within the lobules, sperm develop in structures called seminiferous tubules (FIgure 14.11).

The tightly coiled seminiferous tubules form the bulk of each testis. They are composed of developing sperm cells surrounding a lumen, the hollow center of the tubule, where formed sperm are released into the duct system of the testis. Specifically, from the lumens of the seminiferous tubules, sperm move into the straight tubules (or tubuli recti), and from there into a fine meshwork of tubules called the rete testes. Sperm leave the rete testes, and the testis itself, through the 15 to 20 efferent ductules that cross the tunica albuginea.
Structure of Formed Sperm
Sperm are smaller than most cells in the body; in fact, the volume of a sperm cell is 85,000 times less than that of the female gamete. Approximately 100 to 300 million sperm are produced each day, whereas women typically ovulate only one oocyte per month as is true for most cells in the body, the structure of sperm cells speaks to their function. Sperm have a distinctive head, mid-piece, and tail region (Figure 14.12). A structure called the acrosome covers most of the head of the sperm cell as a “cap” that is filled with lysosomal enzymes important for preparing sperm to participate in fertilization. Tightly packed mitochondria fill the mid-piece of the sperm. ATP produced by these mitochondria will power the flagellum, which extends from the neck and the mid-piece through the tail of the sperm, enabling it to move the entire sperm cell.

To fertilize an egg, sperm must be moved from the seminiferous tubules in the testes, through the epididymis, and—later during ejaculation—along the length of the penis and out into the female reproductive tract.
Epididymis
From the lumen of the seminiferous tubules, the immotile sperm are surrounded by testicular fluid and moved to the epididymis (plural = epididymides), a coiled tube attached to the testis where newly formed sperm continue to mature. Though the epididymis does not take up much room in its tightly coiled state, it would be approximately 6 m (20 feet) long if straightened. Sperm enter the head of the epididymis and are moved along predominantly by the contraction of smooth muscles lining the epididymal tubes. As they are moved along the length of the epididymis, the sperm further mature and acquire the ability to move under their own power. The more mature sperm are then stored in the tail of the epididymis (the final section) until ejaculation occurs.
Duct System and Accessory Glands
During ejaculation, sperm exit the tail of the epididymis and are pushed by smooth muscle contraction to the ductus deferens (also called the vas deferens). The ductus deferens is a thick, muscular tube that is bundled together inside the scrotum with connective tissue, blood vessels, and nerves into a structure called the spermatic cord (Figure 14.10). Because the ductus deferens is physically accessible within the scrotum, surgical sterilization to interrupt sperm delivery can be performed by cutting and sealing a small section of the ductus (vas) deferens. This procedure is called a vasectomy, and it is an effective form of male birth control.
Each ductus deferens extends superiorly into the abdominal cavity through the inguinal canal in the abdominal wall. From here, the ductus deferens continues posteriorly to the pelvic cavity, ending posterior to the bladder where it dilates in a region called the ampulla (meaning “flask”).
Sperm make up only 5 percent of the final volume of semen, the thick, milky fluid that the male ejaculates. The bulk of semen is produced by three critical accessory glands of the male reproductive system: the seminal vesicles, the prostate, and the bulbourethral glands.
As sperm pass through the ampulla of the ductus deferens at ejaculation, they mix with fluid from the associated seminal vesicle (Figure 14.9). The paired seminal vesicles are glands that contribute approximately 60 percent of the semen volume. Seminal vesicle fluid contains large amounts of fructose, which is used by the sperm mitochondria to generate ATP to allow movement through the female reproductive tract.
The fluid, now containing both sperm and seminal vesicle secretions, next moves into the associated ejaculatory duct, a short structure formed from the ampulla of the ductus deferens and the duct of the seminal vesicle. The paired ejaculatory ducts transport the seminal fluid into the next structure, the prostate gland.
The centrally located prostate gland sits anterior to the rectum at the base of the bladder surrounding the prostatic urethra (the portion of the urethra that runs within the prostate). About the size of a walnut, the prostate is formed of both muscular and glandular tissues. It excretes an alkaline, milky fluid to the passing seminal fluid—now called semen—that is critical to first coagulate and then decoagulate the semen following ejaculation. The temporary thickening of semen helps retain it within the female reproductive tract, providing time for sperm to utilize the fructose provided by seminal vesicle secretions. When the semen regains its fluid state, sperm can then pass farther into the female reproductive tract.
The final addition to semen is made by two bulbourethral glands (or Cowper’s glands) that release a thick, salty fluid that lubricates the end of the urethra and the vagina, and helps to clean urine residues from the penile urethra. The fluid from these accessory glands is released after the male becomes sexually aroused, and shortly before the release of the semen. It is therefore sometimes called pre-ejaculate.
The Penis
The penis is the male organ of copulation (sexual intercourse). It is flaccid for non-sexual actions, such as urination, and turgid and rod-like with sexual arousal. When erect, the stiffness of the organ allows it to penetrate into the vagina and deposit semen into the female reproductive tract.
The shaft of the penis surrounds the urethra (Figure 14.13). The shaft is composed of three column-like chambers of erectile tissue that span the length of the shaft. Each of the two larger lateral chambers is called a corpus cavernosum (plural = corpora cavernosa). Together, these make up the bulk of the penis. The corpus spongiosum, which can be felt as a raised ridge on the erect penis, is a smaller chamber that surrounds the spongy, or penile, urethra. The end of the penis, called the glans penis, has a high concentration of nerve endings, resulting in very sensitive skin that influences the likelihood of ejaculation. The skin from the shaft extends down over the glans and forms a collar called the prepuce (or foreskin).

The Female Reproductive System
The female reproductive system produces gametes and reproductive hormones. In addition, the female reproductive system supports the developing fetus and delivers it to the outside world. The female reproductive system is located primarily inside the pelvic cavity. The female gonads are called ovaries and the gamete they produce is called an oocyte.
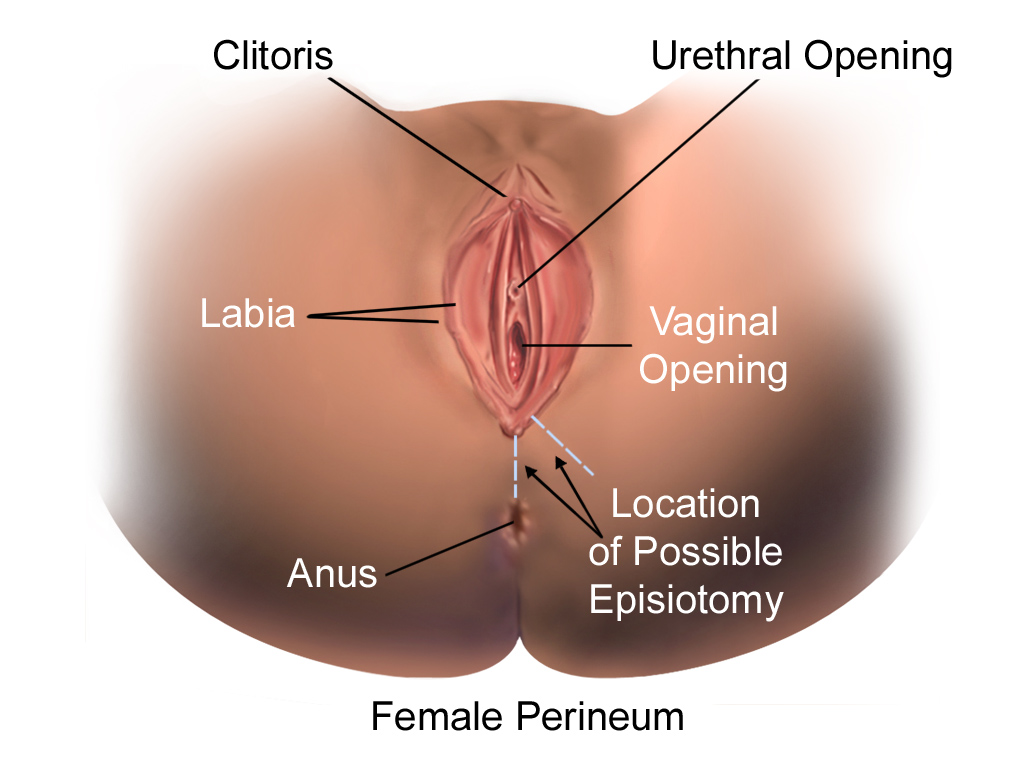
External Female Genitals
The external female reproductive structures are referred to collectively as the vulva and they include:
- The mons pubis is a pad of fat that is located at the anterior, over the pubic bone. After puberty, it becomes covered in pubic hair.
- The labia majora (labia = “lips”; majora = “larger”) are folds of hair-covered skin that begin just posterior to the mons pubis.
- The labia minora (labia = “lips”; minora = “smaller”) is thinner and more pigmented and extends medially to the labia majora.
- Although they naturally vary in shape and size from woman to woman, the labia minora serve to protect the female urethra and the entrance to the female reproductive tract.
- The superior, anterior portions of the labia minora come together to encircle the clitoris (or glans clitoris), an organ that originates from the same cells as the glans penis and has abundant nerves that make it important in sexual sensation and orgasm. The hymen is a thin membrane that sometimes partially covers the entrance to the vagina.
- The vaginal opening is located between the opening of the urethra and the anus (Figure 14.14).
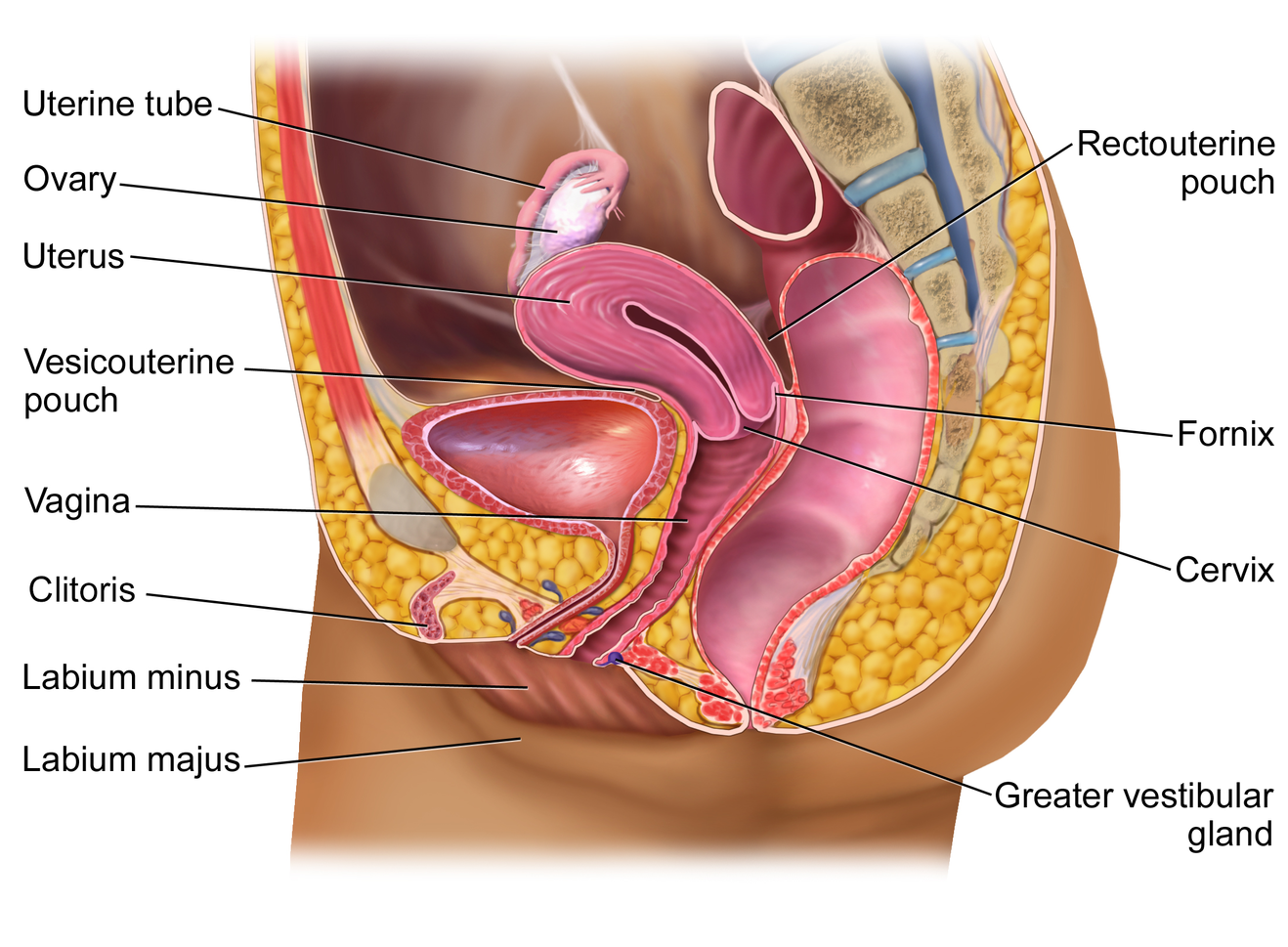
Internal Female Reproductive Organs
Ovaries
The ovaries are the female gonads. There are two, one at each entrance to the uterine (fallopian) tube (Figure 14.15). They are each about 2 to 3 cm in length, about the size of an almond. The ovaries are located within the pelvic cavity. The ovary itself is attached to the uterus via the ovarian ligament. Oocytes, surrounded by supporting cells, develop within each ovary. This grouping of an oocyte and its supporting cells is called a follicle.
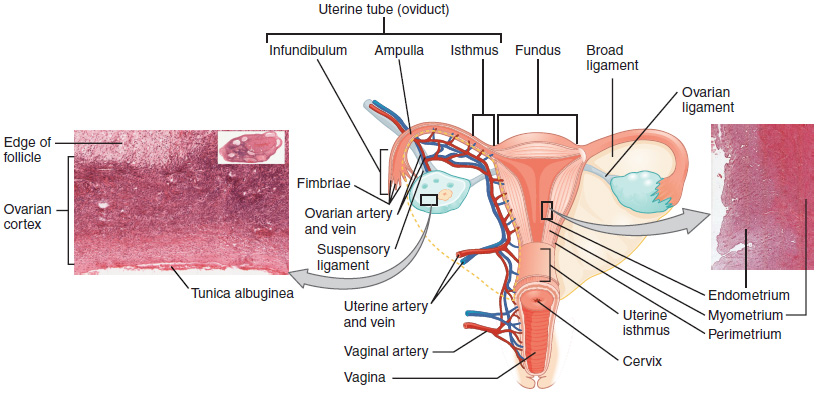
The Uterine Tubes
The uterine (fallopian) tubes are the conduit of the oocyte from the ovary to the uterus. Each of the two fallopian tubes is close to, but not directly connected to, the ovary. The uterine tubes are lined with ciliated cells that beat in the direction of the uterus, producing a current that will be critical to moving the oocyte towards the uterus. The wide distal infundibulum of the tube flares out with slender, finger-like projections called fimbriae, which envelop the ovary. The middle region of the tube, called the ampulla, is where fertilization often occurs. The isthmus is the narrow medial end of each uterine tube that is connected to the uterus.
The Uterus and Cervix
The uterus is the muscular organ that nourishes and supports the growing embryo. Its average size is approximately 5 cm wide by 7 cm long and it has three sections (Figure 14.16). The portion of the uterus superior to the opening of the uterine tubes is called the fundus. The middle section of the uterus is called the body of uterus (or corpus). The cervix is the narrow inferior portion of the uterus that projects into the vagina. The cervix produces mucus secretions that become thin and stringy under the influence of high systemic plasma estrogen concentrations, and these secretions can facilitate sperm movement through the reproductive tract.
The wall of the uterus is made up of three layers:
- Perimetrium: the most superficial layer and serous membrane.
- Myometrium: a thick layer of smooth muscle responsible for uterine contractions.
- Endometrium: the innermost layer containing a connective tissue lining covered by epithelial tissue that lines the lumen. It provides the site of implantation for a fertilized egg, and sheds during menstruation if no egg is fertilized.
Vagina
The vagina is a muscular canal (approximately 10 cm long) that is the entrance to the reproductive tract. It also serves as the exit from the uterus during menses and childbirth. The outer walls of the anterior and posterior vagina are columns with ridges. The superior fornix meets the uterine cervix.
The walls of the vagina are lined with:
- An outer, fibrous adventitia
- A middle layer of smooth muscle
- An inner mucous membrane with transverse folds called rugae.
Together, the middle and inner layers allow the expansion of the vagina to accommodate intercourse and childbirth. The thin, perforated hymen can partially surround the opening to the vaginal orifice. The lesser and greater vestibular glands (located near the clitoris and external opening of the vagina, respectively) secrete mucus, which keeps the vestibular area moist.
Development of Oocytes
Gametogenesis in females is called oogenesis. The process begins with the ovarian stem cells, or oogonia. Oogonia are formed during fetal development, and divide via mitosis, much like spermatogonia in the testis. Unlike spermatogonia, however, oogonia form primary oocytes in the fetal ovary prior to birth. These primary oocytes are then arrested in this stage of meiosis I, only to resume it years later, beginning at puberty and continuing until the woman is near menopause (the cessation of a woman’s reproductive functions). The number of primary oocytes present in the ovaries declines from one to two million in an infant, to approximately 400,000 at puberty, to zero by the end of menopause.
The initiation of ovulation—the release of an oocyte from the ovary—marks the transition from puberty into reproductive maturity for women. From then on, throughout a woman’s reproductive years, ovulation occurs approximately once every 28 days. Just prior to ovulation, a surge of luteinizing hormone triggers the resumption of meiosis in a primary oocyte. This initiates the transition from primary to secondary oocyte. However, this cell division does not result in two identical cells. Instead, the cytoplasm is divided unequally, and one daughter cell is much larger than the other. This larger cell, the secondary oocyte, eventually leaves the ovary during ovulation. The smaller cell, called the first polar body, may or may not complete meiosis and produce second polar bodies; in either case, it eventually disintegrates. Therefore, even though oogenesis produces up to four cells, only one survives.
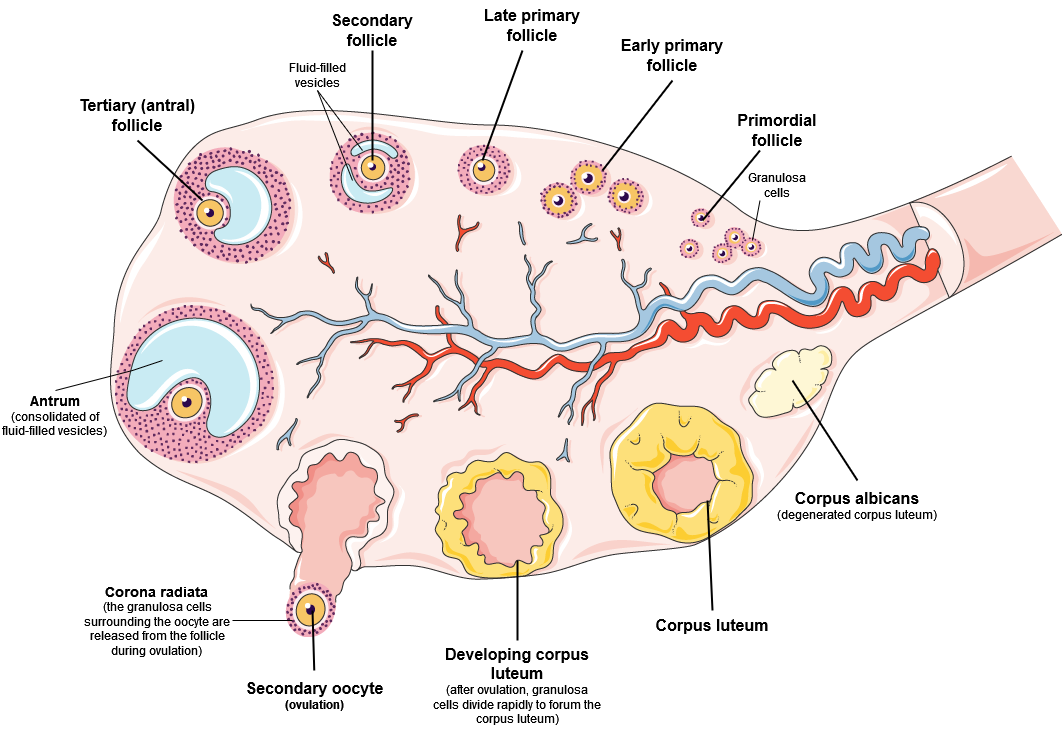
Ovarian follicles are oocytes and their supporting cells. They grow and develop in a process called folliculogenesis, which typically leads to ovulation of one follicle approximately every 28 days, along with death to multiple other follicles. Follicles progress from primordial, to primary, to secondary and tertiary (antral, or mature) stages prior to ovulation—with the oocyte inside the follicle remaining as a primary oocyte until right before ovulation.
Folliculogenesis begins with follicles in a resting state. These small primordial follicles are present in newborn females and are the prevailing follicle type in the adult ovary (Figure 14.17). Primordial follicles have only a single flat layer of support cells, called granulosa cells, that surround the oocyte, and they can stay in this resting state for years—some until right before menopause. After puberty, a few primordial follicles will respond to a recruitment signal each day, and will join a pool of immature growing follicles called primary follicles. Primary follicles start with a single layer of granulosa cells, but the granulosa cells then become active and transition from a flat or squamous shape to a rounded, cuboidal shape as they increase in size and proliferate. As the granulosa cells divide, the follicles—now called secondary follicles – increase in diameter, adding a new outer layer of connective tissue, blood vessels, and theca cells—cells that work with the granulosa cells to produce estrogens.
Within the growing secondary follicle, the primary oocyte now secretes a thin acellular membrane called the zona pellucida that will play a critical role in fertilization. A thick fluid, called follicular fluid, that has formed between the granulosa cells also begins to collect into one large pool, or antrum. Follicles in which the antrum has become large and fully formed are considered tertiary follicles (or antral follicles). Several follicles reach the tertiary stage at the same time, and most of these will undergo atresia (death). The one that does not die will continue to grow and develop until ovulation, when it will expel its secondary oocyte surrounded by several layers of granulosa cells from the ovary. Keep in mind that most follicles don’t make it to this point. In fact, roughly 99 percent of the follicles in the ovary will undergo atresia, which can occur at any stage of folliculogenesis.
Once the secondary oocyte has been ovulated, there is a change in the follicular support cells the formed the tertiary follicle. The collapsed follicle is transformed into a new endocrine structure called the corpus luteum, a term meaning “yellowish body”. The corpus luteum produces a hormone (progesterone) that is critical for the establishment and maintenance of pregnancy. If pregnancy does not occur within 10 to 12 days, the corpus luteum will stop secreting progesterone and degrade into the corpus albicans, a nonfunctional “whitish body” that will disintegrate in the ovary over a period of several months (Figure 14.17).
Ovulation
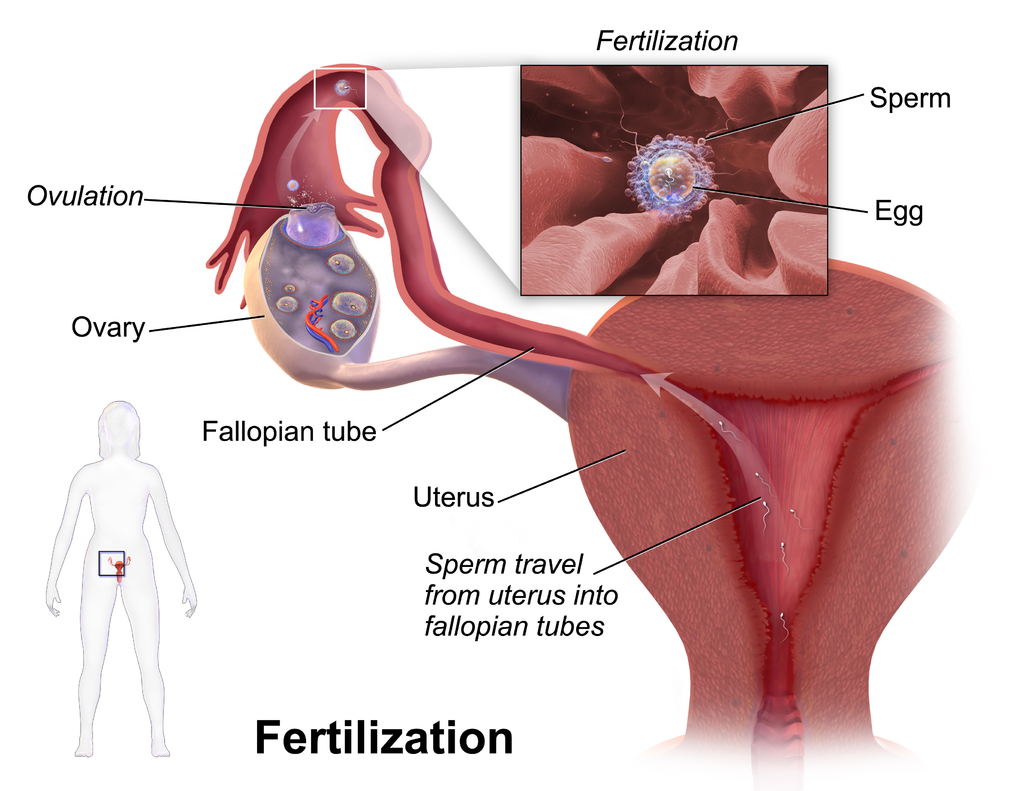
Following ovulation, the uterine tube receives the oocyte. Oocytes lack flagella, and therefore cannot move on their own. High concentrations of estrogen that occur around the time of ovulation induce contractions of the smooth muscle along the length of the uterine tube. These contractions occur every 4 to 8 seconds, causing the oocyte to flow towards the uterus, through the coordinated beating of the cilia that line the outside and lumen of the length of the Fallopian tube which pulls the oocyte into the interior of the tube. Once inside, the muscular contractions and beating cilia move the oocyte slowly toward the uterus. When fertilization does occur, sperm typically meet the egg while it is still moving through the ampulla (Figure 14.18).
Unless otherwise indicated, this chapter contains material adapted from chapters 17 and 19 in Anatomy, Physiology, and Medical Language by NSCC, Kimberlee Carter, Marie Rutherford, and Douglas College Biology Department as well as chapters 25 and 27 in Anatomy and Physiology (on OpenStax), by Betts, et al. and is used under a a CC BY 4.0 international license. Download and access OpenStax Anatomy and Physiology for free at https://openstax.org/books/anatomy-and-physiology-2e/

{kind=link}
{kind=link}
{kind=link}
{kind=link}