
Lab 3: Tissues II and Integumentary system
Connective, muscle, and nervous tissues; the integumentary system
When you are prepared for the Test on Week 3 Learning Objectives in Week 4, you will be able to:
- Review from Week 2: Differentiate between the general functions of epithelial, connective, muscle, and nervous tissues.
- Review from Week 2: Identify epithelial tissue types, their features, and their basic functions.
- Identify connective tissue types, their features, and their basic functions.
- Identify muscle tissue types.
- Identify nervous tissue.
- Identify layers of the skin, layers within the epidermis, and their functions.
- Identify accessory structures in the skin and explain their functions.
Last week we looked at the four different categories of tissue in the body: epithelial, connective, muscle, and nervous. We then observed different types of epithelial tissue and learned how to differentiate between them. This week we will look at examples of the remaining three tissue categories and learn how to differentiate between specific types of connective tissue and muscle tissue. We will then look at the first organ system of the class, the Integumentary system. This consists of the skin and its accessory structures.
Tissues II
Connective tissue
General structure of connective tissue
As may be obvious from its name, one of the major functions of connective tissue is to connect tissues and organs. Unlike epithelial tissue, which is composed of cells closely packed with little or no extracellular space in between, connective tissue cells are dispersed in a matrix. The matrix usually includes a large amount of extracellular material produced by the connective tissue cells that are embedded within it. The matrix plays a major role in the functioning of this tissue. The major component of the matrix is a ground substance often crisscrossed by protein fibers. This ground substance is usually a fluid, but it can also be mineralized and solid, as in bones. Connective tissues come in a vast variety of forms, yet they typically have in common three characteristic components: cells, large amounts of amorphous ground substance, and protein fibers. The amount and structure of each component correlates with the function of the tissue, from the rigid ground substance in bones supporting the body to the inclusion of specialized cells; for example, a phagocytic cell that engulfs pathogens and also rids tissue of cellular debris.
Functions of Connective Tissues
Connective tissues perform many functions in the body, but most importantly, they support and connect other tissues; from the connective tissue sheath that surrounds muscle cells, to the tendons that attach muscles to bones, and to the skeleton that supports the positions of the body. Protection is another major function of connective tissue, in the form of fibrous capsules and bones that protect delicate organs and, of course, the skeletal system. Specialized cells in connective tissue defend the body from microorganisms that enter the body. Transport of fluid, nutrients, waste, and chemical messengers is ensured by specialized fluid connective tissues, such as blood and lymph. Adipose cells store surplus energy in the form of fat and contribute to the thermal insulation of the body.
Classification of Connective Tissue
The three broad categories of connective tissue are classified according to the characteristics of their ground substance and the types of fibers found within the matrix (Table 3.1). Connective tissue proper includes loose connective tissue and dense connective tissue. Both tissues have a variety of cell types and protein fibers suspended in a viscous ground substance. Dense connective tissue is reinforced by bundles of fibers that provide tensile strength, elasticity, and protection. In loose connective tissue, the fibers are loosely organized, leaving large spaces in between. Supportive connective tissue – bone and cartilage – provide structure and strength to the body and protect soft tissues. A few distinct cell types and densely packed fibers in a matrix characterize these tissues. In bone, the matrix is rigid and described as calcified because of the deposited calcium salts. In fluid connective tissue – lymph and blood – various specialized cells circulate in a watery fluid containing salts, nutrients, and dissolved proteins.
| Connective tissue proper | Supportive connective tissue | Fluid connective tissue |
|---|---|---|
|
|
Blood |
|
|
Lymph |
Connective Tissue Proper
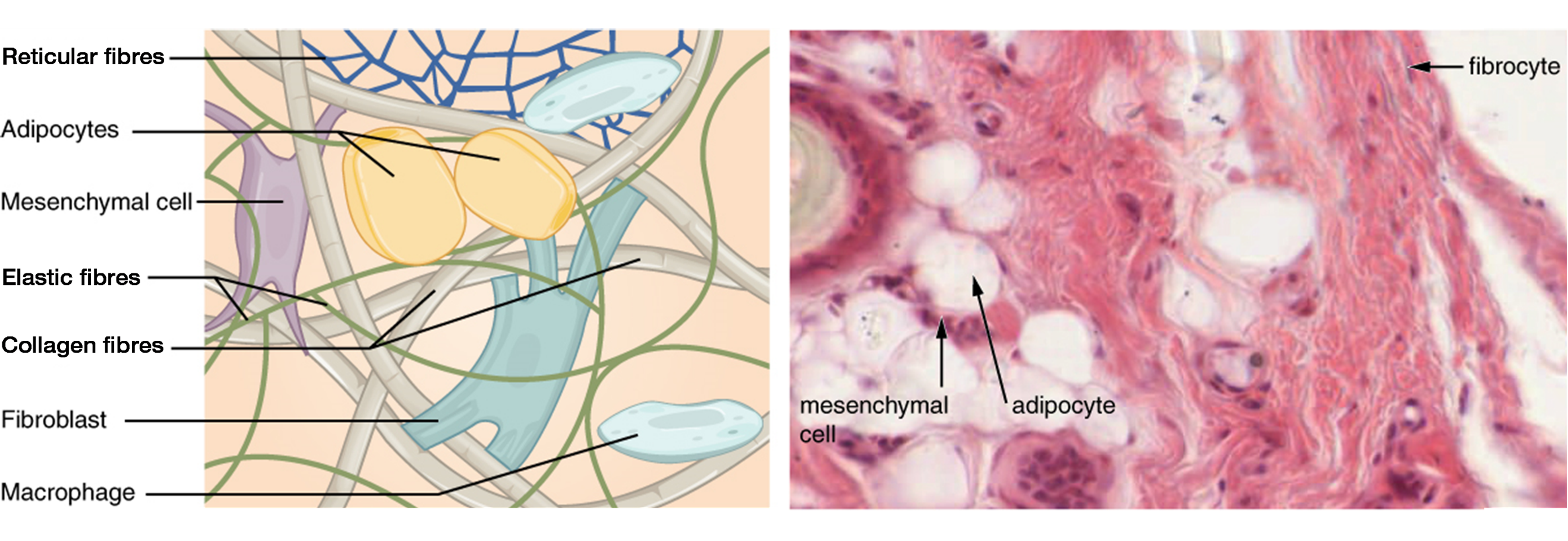
The most abundant cell in connective tissue proper is the fibroblast. Fibroblasts secrete a viscous ground substance that, with embedded fibrous proteins, forms the extra-cellular matrix (Figure 3.1). Fibrocytes, along with adipocytes, and mesenchymal cells, are fixed cells, which means they remain within the connective tissue. Adipocytes are cells that store lipids as droplets that fill most of the cytoplasm. The mesenchymal cell is a multipotent adult stem cell. These cells can differentiate into any type of connective tissue cells needed for repair and healing of damaged tissue. Other cells move in and out of the connective tissue in response to chemical signals. Macrophages, mast cells, lymphocytes, plasma cells, and phagocytic cells are found in connective tissue proper but are actually part of the immune system protecting the body.
Three main types of protein fibers are secreted by fibroblasts: collagen fibers, elastic fibers, and reticular fibers. Collagen fibers, while flexible, have great tensile strength, resist stretching, and give ligaments and tendons their characteristic resilience and strength. Elastic fibers, after being stretched or compressed will return to its original shape. Reticular fibers are narrow and are arrayed in a branching network. They are found throughout the body, but are most abundant in the reticular tissue of soft organs, such as liver and spleen, where they anchor and provide structural support to the the functional cells, blood vessels, and nerves of the organ. All of these fiber types are embedded in ground substance, a clear, viscous, colorless matrix made of polysaccharides and proteins, forming the extracellular matrix of connective tissue proper (Figure 3.1).
Loose Connective Tissue
Loose connective tissue is found between many organs where it acts both to absorb shock and bind tissues together. It allows water, salts, and various nutrients to diffuse through to adjacent or imbedded cells and tissues.
Areolar tissue shows little specialization. It contains all the cell types and fibers previously described and is distributed in a random, web-like fashion. It fills the spaces between muscle fibers, surrounds blood and lymph vessels, and supports organs in the abdominal cavity. Areolar tissue underlies most epithelia and represents the connective tissue component of epithelial membranes, which are described further in a later section.
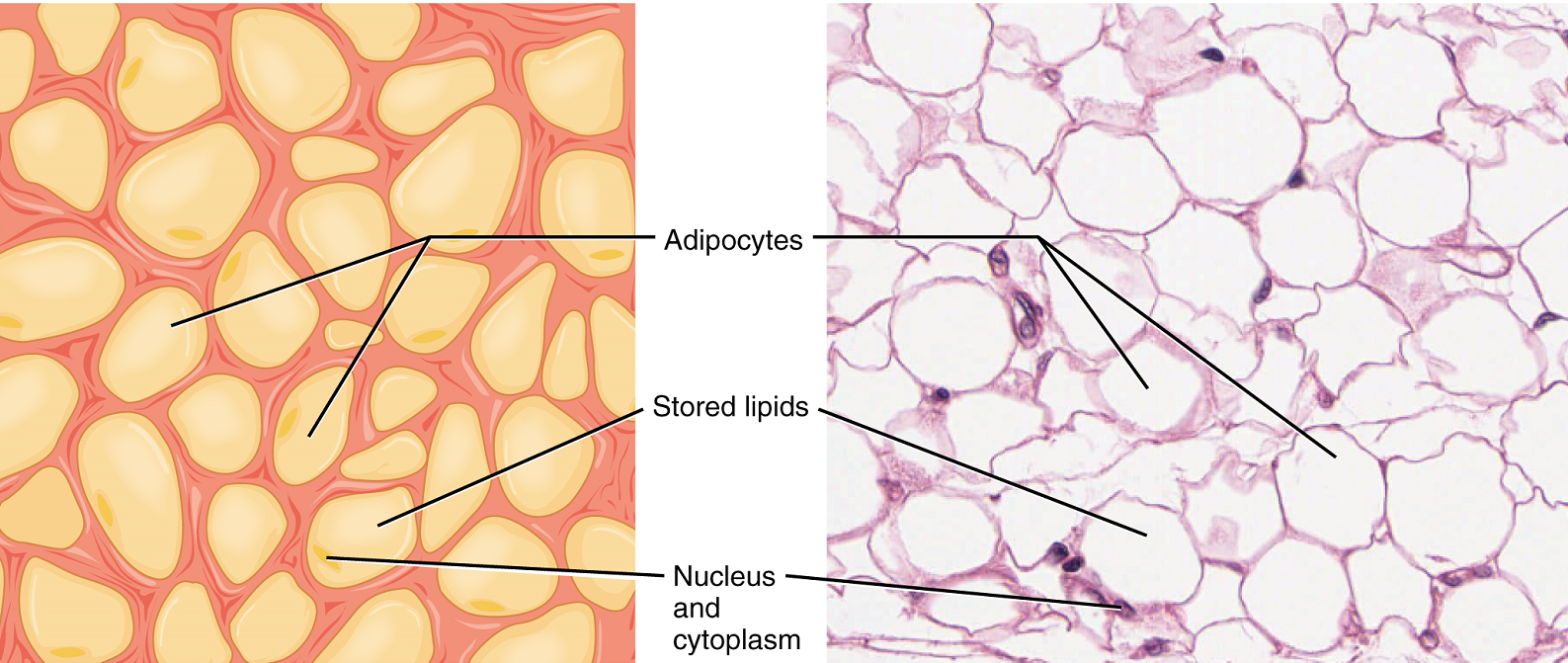
Adipose tissue consists mostly of fat storage cells, with little extracellular matrix (Figure 3.2). A large number of capillaries allow rapid storage and mobilization of lipid molecules. Fat contributes mostly to lipid storage and can serve as insulation from cold temperatures and mechanical injuries.
Reticular tissue is a mesh-like, supportive framework for soft organs such as lymphatic tissue, the spleen, and the liver (Figure 3.3). Reticular cells produce the reticular fibers that form the network onto which other cells attach. It derives its name from the Latin reticulus, which means “little net.”
Dense Connective Tissue
Dense connective tissue contains more collagen fibers than does loose connective tissue. As a consequence, it displays greater resistance to stretching. There are two major categories of dense connective tissue: regular and irregular. Dense regular connective tissue fibers are parallel to each other, enhancing tensile strength and resistance to stretching in the direction of the fiber orientations. Ligaments and tendons are made of dense regular connective tissue, but in ligaments not all fibers are parallel. Dense regular elastic connective tissue contains elastin fibers in addition to collagen fibers, which allows the ligament to return to its original length after stretching. The ligaments in the vocal folds and between the vertebrae in the vertebral column are elastic.
In dense irregular connective tissue, the direction of fibers is random. This arrangement gives the tissue greater strength in all directions and less strength in one particular direction. In some tissues, fibers crisscross and form a mesh. In other tissues, stretching in several directions is achieved by alternating layers where fibers run in the same orientation in each layer, and it is the layers themselves that are stacked at an angle. The dermis of the skin is an example of dense irregular connective tissue rich in collagen fibers. Dense irregular elastic connective tissue give arterial walls the strength and the ability to regain original shape after stretching (Figure 3.4).
Supportive Connective Tissues
Two major forms of supportive connective tissue, cartilage and bone, allow the body to maintain its posture and protect internal organs.
Cartilage
The distinctive appearance of cartilage is due to polysaccharides, which bind with ground substance proteins to form the extracellular matrix. Embedded within the cartilage matrix are chondrocytes, or cartilage cells, and the space they occupy are called lacunae (singular = lacuna). Cartilaginous tissue is avascular, thus all nutrients need to diffuse through the matrix to reach the chondrocytes. This is a factor contributing to the very slow healing of cartilaginous tissues.
The three main types of cartilage tissue are hyaline cartilage, fibrocartilage, and elastic cartilage (Figure 3.5). Hyaline cartilage, the most common type of cartilage in the body, contains short and dispersed collagen fibers in the matrix. Both strong and flexible, the hyaline cartilage is found in the rib cage and nose and covers bones where they meet to form moveable joints. It makes up a template of the embryonic skeleton before bone formation. A plate of hyaline cartilage at the ends of bone allows continued growth until adulthood. Fibrocartilage is tough because it has thick bundles of collagen fibers dispersed through its matrix. The knee and jaw joints and the intervertebral discs are examples of fibrocartilage. Elastic cartilage contains elastic fibers as well as collagen. This tissue gives rigid support as well as elasticity. Tug gently at your ear lobes, and notice that the lobes return to their initial shape. The external ear contains elastic cartilage.
Bone
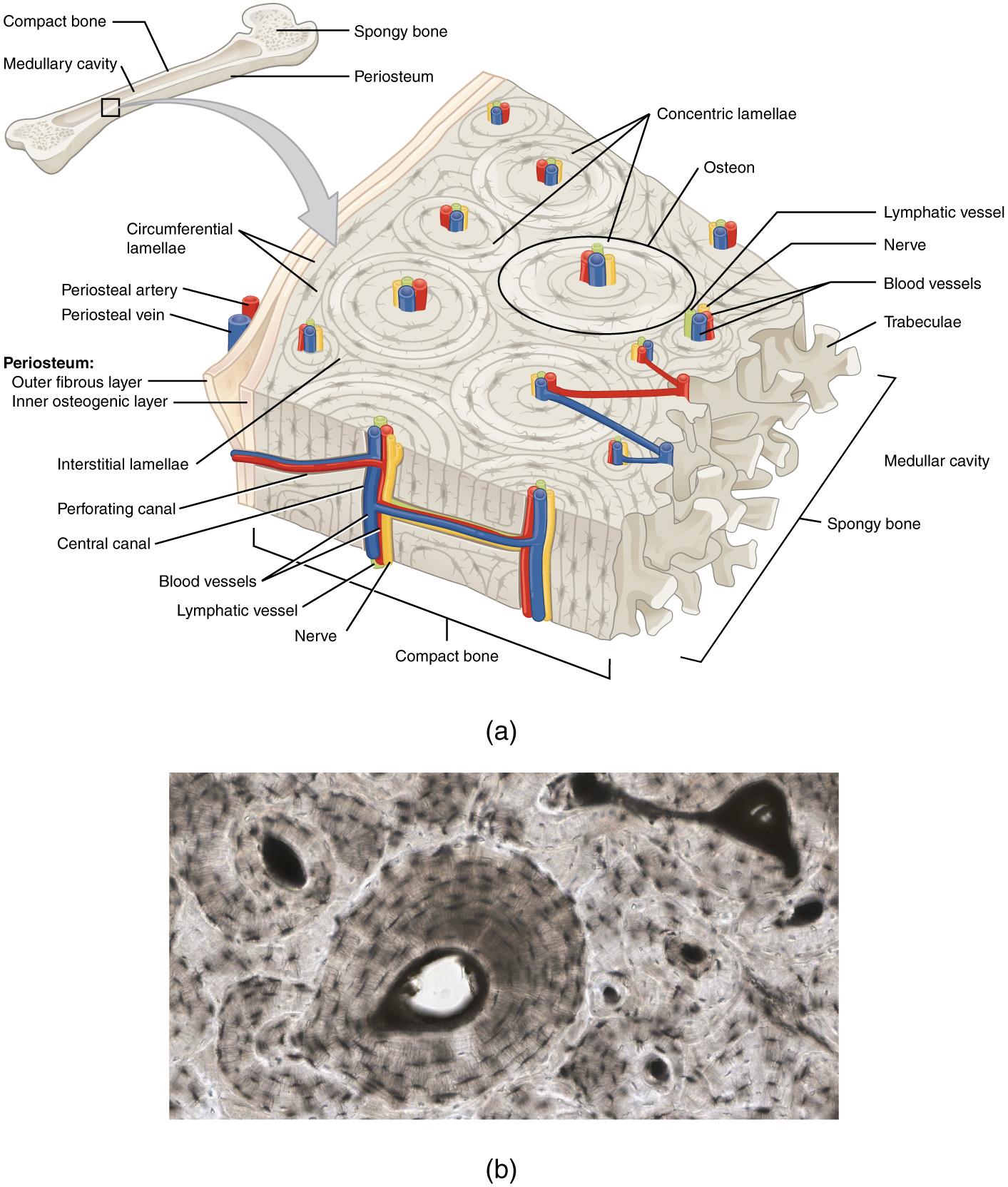
Bone is the hardest connective tissue. It provides protection to internal organs and supports the body. Bone’s rigid extracellular matrix contains mostly collagen fibers embedded in a mineralized ground substance containing hydroxyapatite, a form of calcium phosphate. Both components of the matrix, organic and inorganic, contribute to the unusual properties of bone. Without collagen, bones would be brittle and shatter easily. Without mineral crystals, bones would flex and provide little support. Osteocytes, bone cells similar to chondrocytes, are located within lacunae. Bone is a highly vascularized tissue. Unlike cartilage, bone tissue can recover from injuries in a relatively short time.
Compact bone tissue is solid and has great structural strength. It is found on the outside of bones and reinforcing the shafts of long bones. The histology of a transverse section of compact bone shows a typical arrangement of osteocytes in concentric circles around a central canal (Figure 3.6). Spongy bone looks like a sponge under the microscope and contains empty spaces between trabeculae, or arches of bone proper. It is lighter than compact bone and found in the interior of some bones and at the end of long bones.
Fluid Connective Tissue
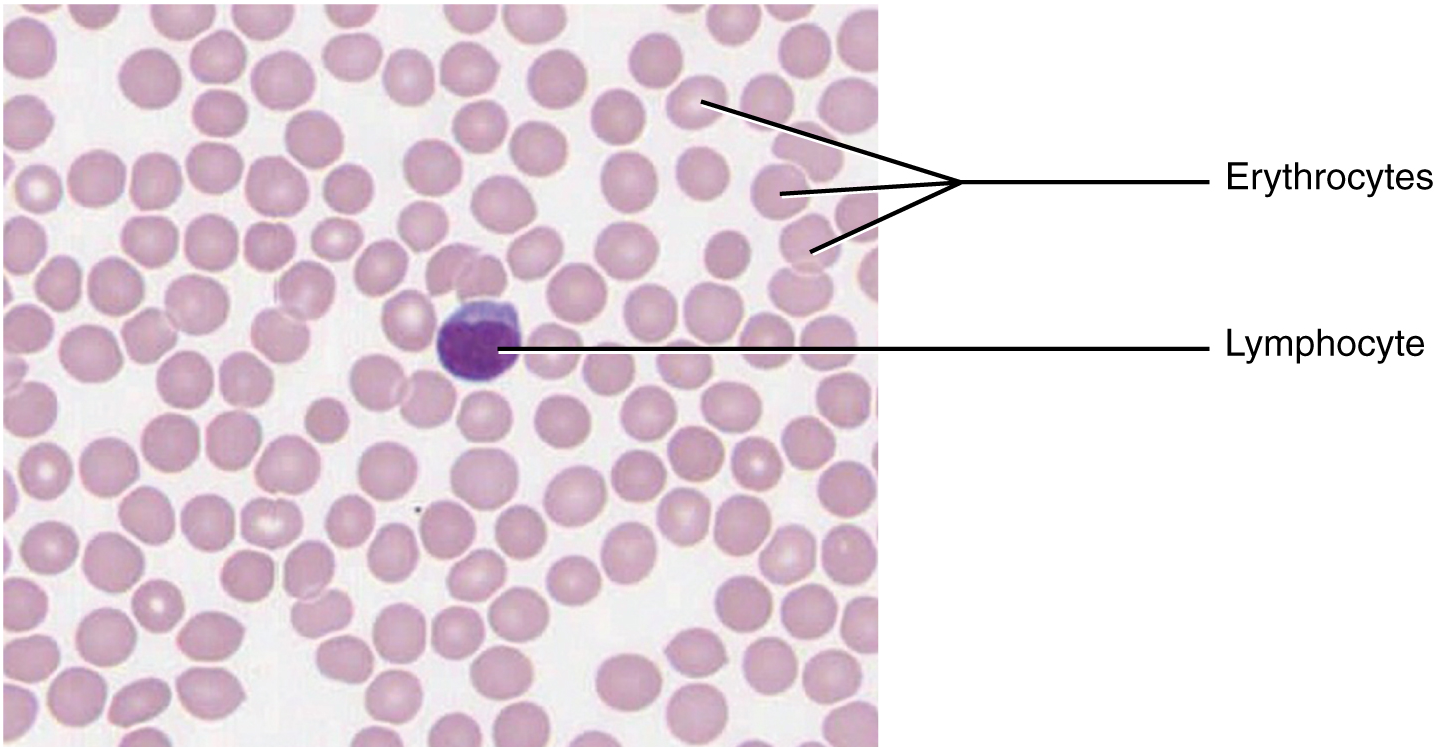
Blood and lymph are fluid connective tissues. Cells circulate in a liquid extracellular matrix. The formed elements circulating in blood are all derived from hematopoietic stem cells located in bone marrow. In blood, erythrocytes (red blood cells) transport oxygen and some carbon dioxide (Figure 3.7). Leukocytes (white blood cells) are responsible for defending against potentially harmful microorganisms or molecules. Platelets are cell fragments involved in blood clotting. Nutrients, salts, and wastes are dissolved in the liquid matrix and transported through the body.
Lymph contains a liquid matrix and white blood cells. Lymphatic capillaries are extremely permeable, allowing larger molecules and excess fluid from interstitial spaces to enter the lymphatic vessels. Lymph drains into blood vessels, delivering molecules to the blood that could not otherwise directly enter the bloodstream. In this way, specialized lymphatic capillaries transport absorbed fats away from the intestine and deliver these molecules to the blood.
Muscle Tissue
Muscle tissue is characterized by properties that allow movement. Muscle cells are excitable; they respond to a stimulus. They are contractile, meaning they can shorten and generate a pulling force. When attached between two movable objects, in other words, bones, contractions of the muscles cause the bones to move. Some muscle movement is voluntary, which means it is under conscious control. For example, a person decides to open a book and read a chapter on anatomy. Other movements are involuntary, meaning they are not under conscious control, such as the contraction of your pupil in bright light. Muscle tissue is classified into three types according to structure and function: skeletal, cardiac, and smooth (Table 3.2).
| Tissue | Histology | Function | Location |
|---|---|---|---|
| Skeletal | Long cylindrical fiber; striated; many peripherally-located nuclei | Voluntary movement; thermogenesis; organ protection | Attached to bones; found around entrance points to body (e.g. mouth, anus) |
| Cardiac | Short, branched fibers; striated; single central nucleus | Contracts to pump blood | Heart walls |
| Smooth | Short, spindle-shaped fibers; no evident striation; single nucleus | Involuntary movement; moves material through digestive tract and ducts; regulates blood flow in arteries | Walls of major organs and passageways |
Skeletal muscle
Skeletal muscle tissue is attached to bones and its contraction makes possible locomotion, facial expressions, posture, and other voluntary movements of the body. Forty percent of your body mass is made up of skeletal muscle. Skeletal muscles generate heat as a byproduct of their contraction and thus participate in thermal homeostasis. Shivering is an involuntary contraction of skeletal muscles in response to perceived lower than normal body temperature.
The muscle cells (muscle fibers) and their numbers remain relatively constant throughout life. Skeletal muscle tissue is arranged in bundles surrounded by connective tissue. Under the light microscope, muscle cells appear striated with many nuclei squeezed along the membranes (Figure 3.8a). The striation is due to the regular alternation of the contractile proteins actin and myosin, along with the structural proteins that couple the contractile proteins to connective tissues. The cells are multinucleated as a result of the fusion of the many myoblasts that fuse to form each long muscle fiber.
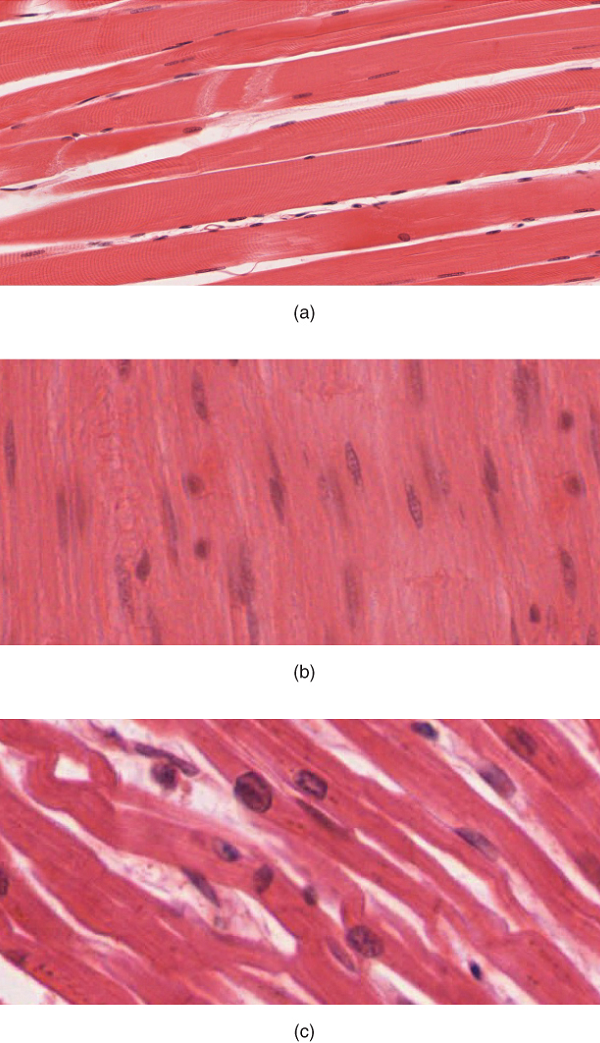
Cardiac muscle
Cardiac muscle tissue forms the contractile walls of the heart. The cells of cardiac muscle, known as cardiomyocytes, also appear striated under the microscope. Unlike skeletal muscle fibers, cardiomyocytes are single cells typically with a single centrally located nucleus (Figure 3.8c).
A principal characteristic of cardiomyocytes is that they contract on their own intrinsic rhythms without any external stimulation. Cardiomyocytes attach to one another with specialized cell junctions called intercalated discs that contain gap junctions. Attached cells form long, branching cardiac muscle fibers and the ions carrying a contraction signal flow rapidly from cell to cell through the gap junctions, which allows the cells to synchronize their actions. The cardiac muscle pumps blood through the body and is under involuntary control.
Smooth muscle
Smooth muscle tissue contraction is responsible for involuntary movements in the internal organs. It forms the contractile component of the digestive, urinary, and reproductive systems as well as the airways and arteries. Each cell is spindle shaped with a single nucleus and no visible striations (Figure 3.8b).
Nervous Tissue
Nervous tissue is characterized as being excitable and capable of sending and receiving electrochemical signals that provide the body with information. Two main classes of cells make up nervous tissue: the neuron and neuroglia . Neurons propagate information via electrochemical impulses, called action potentials, which are biochemically linked to the release of chemical signals. Neuroglia play an essential role in supporting neurons and modulating their information propagation. The presence of the nervous tissue throughout the body and its organization allow it to receive, integrate and provide information to the entire body. This ensures that appropriate responses can occur among all body systems within an intact organism, both under normal conditions as well as during times of stress.
Neurons display distinctive morphology, well suited to their role as conducting cells, with three main parts (Figure 3.9). The cell body includes most of the cytoplasm, the organelles, and the nucleus. Dendrites branch off the cell body and appear as thin extensions. A long “tail,” the axon, extends from the neuron body and can be wrapped in an insulating layer known as myelin, which is formed by accessory cells. The synapse is the gap between nerve cells, or between a nerve cell and its target, for example, a muscle or a gland, across which the impulse is transmitted by chemical compounds known as neurotransmitters.
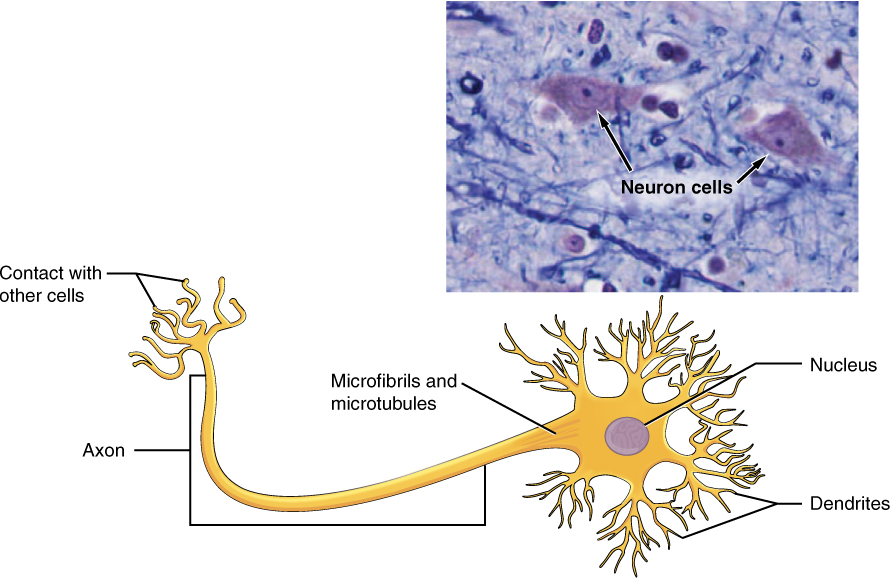
Integumentary System
Although you may not typically think of the skin as an organ, it is in fact made of tissues that work together as a single structure to perform unique and critical functions. The skin and its accessory structures make up the integumentary system, which provides the body with overall protection. The skin is made of multiple layers of cells and tissues, which are held to underlying structures by connective tissue (Figure 3.10). The superficial layer, known as the epidermis, is composed primarily of tightly pack epithelial cells. The deeper layer of skin, the dermis, is well vascularized (has numerous blood vessels) and is where several accessory structures, such as hair follicles, sweat glands, and oil glands, can be found. It also has numerous sensory, and autonomic and sympathetic nerve fibers ensuring communication to and from the brain. Below the dermis lies the superficial fascia, or hypodermis, which is not technically part of the skin, but serves to connect the skin to the underlying fascia. This layer is mainly composed of loose, areolar and adipose connective tissues.
The skin and accessory structures perform a variety of essential functions, such as protecting the body from invasion by microorganisms, chemicals, and other environmental factors; preventing dehydration; acting as a sensory organ; modulating body temperature and electrolyte balance; and synthesizing vitamin D. The underlying hypodermis has important roles in storing fats, forming a “cushion” over underlying structures, and providing insulation from cold temperatures. The skin protects the rest of the body from the basic elements of nature such as wind, water, and UV sunlight. It acts as a protective barrier against water loss, due to the presence of layers of
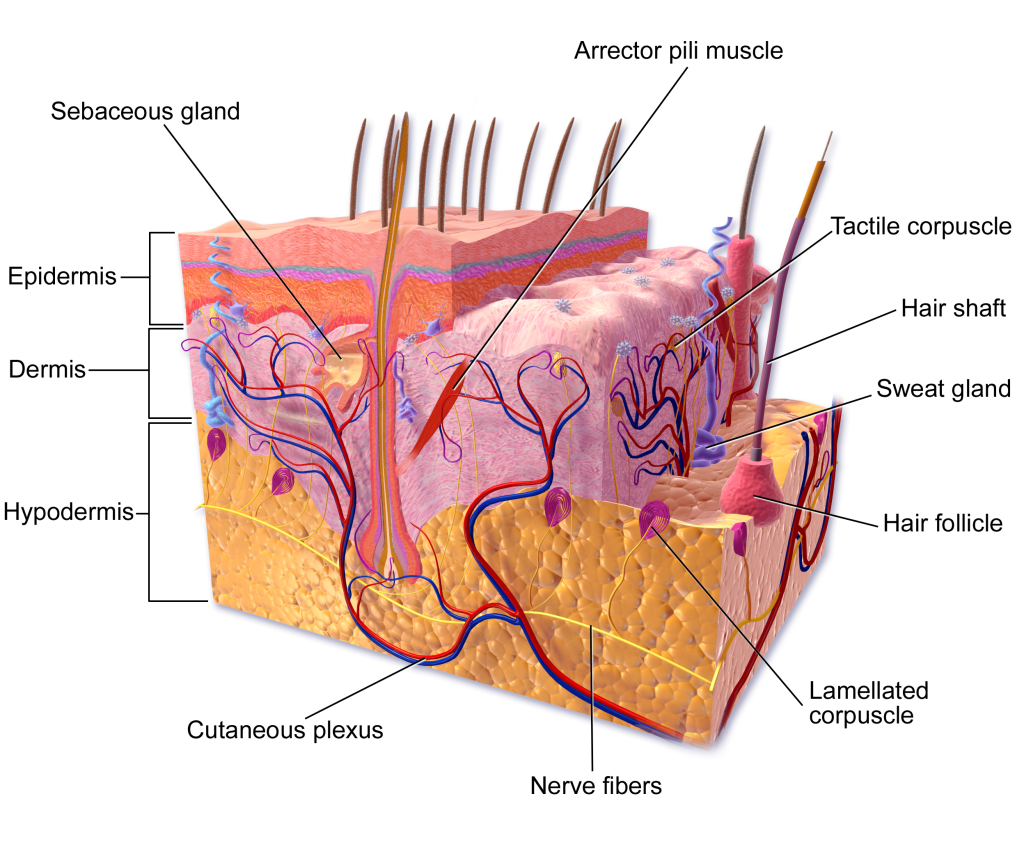
The Epidermis
The epidermis is composed of keratinized, stratified squamous epithelium. It is made of four or five layers, depending on its location in the body. It does not have any blood vessels within it (i.e., it is avascular). Skin that has four epidermal layers is referred to as “thin skin.” From deep to superficial, these layers are the stratum basale, stratum spinosum, stratum granulosum, and stratum corneum. Most of the skin can be classified as thin skin. “Thick skin” is found only on the palms of the hands and the soles of the feet. It has a fifth layer, called the stratum lucidum, located between the stratum corneum and the stratum granulosum (Figure 3.11).
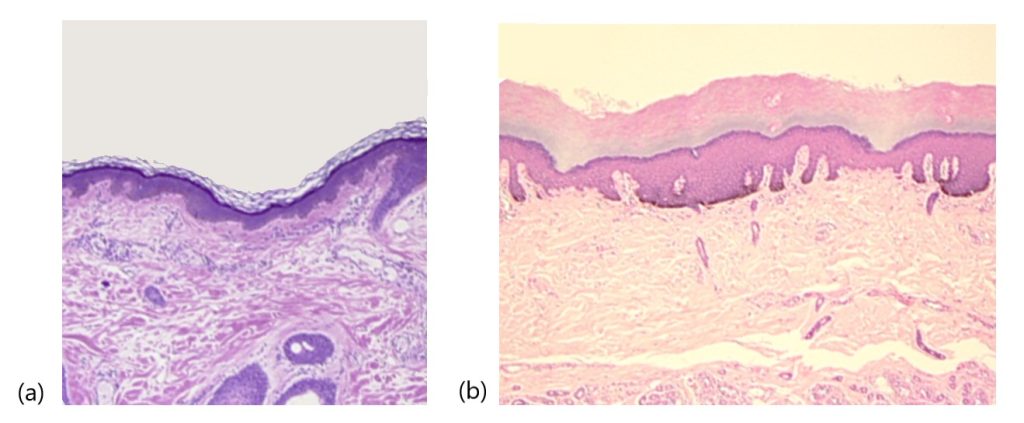
The majority of cells in the epidermis are called keratinocytes. A keratinocyte is a cell that manufactures and stores the protein keratin. Keratin is an intracellular fibrous protein that gives hair, nails, and skin their hardness and water-resistant properties. The keratinocytes in the stratum corneum are dead and regularly slough away, being replaced by cells from the deeper layers.
Stratum Basale
The stratum basale is the deepest epidermal layer and attaches the epidermis to the basal lamina, below which lie the layers of the dermis (Figure 3.12). A finger-like projection, or fold, known as the dermal papilla (plural = dermal papillae) is found in the superficial portion of the dermis. Dermal papillae increase the strength of the connection between the epidermis and dermis; the greater the surface area conveyed by folding, the stronger the connections made . In a growing fetus, fingerprints form where the cells of the stratum basale meet the papillae of the underlying dermal layer (papillary layer), resulting in the formation of the ridges on your fingers that you recognize as fingerprints. Fingerprints are unique to each individual and are used for forensic analyses because the patterns do not change with the growth and aging processes.
The stratum basale is a single layer of cells primarily made of basal cells. A basal cell is a cuboidal-shaped stem cell that is a precursor of the keratinocytes of the epidermis. All of the keratinocytes are produced from this single layer of cells, which are constantly going through mitosis to produce new cells. As new cells are formed, the existing cells are pushed superficially away from the stratum basale. Two other cell types are found dispersed among the basal cells in the stratum basale. The first is a tactile cell (Merkel cell), which functions as a receptor and is responsible for stimulating sensory nerves that the brain perceives as touch. These cells are especially abundant on the surfaces of the hands and feet. The second is a melanocyte, a cell that produces the pigment melanin. Melanin gives hair and skin its color, and also helps protect the living cells of the epidermis from ultraviolet (UV) radiation damage.
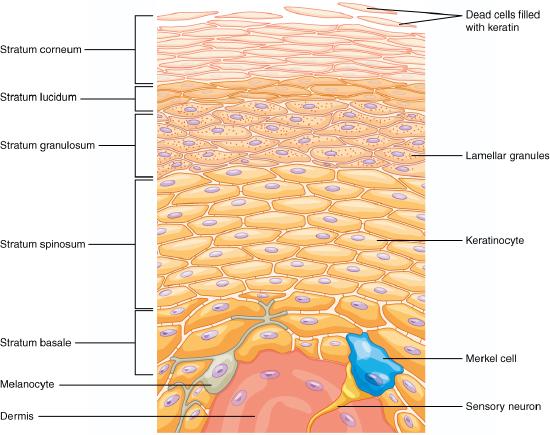
Stratum Spinosum
As the name suggests, the stratum spinosum is spiny in appearance due to the protruding cell processes that join the cells via an anchoring junction common in epithelial tissues called a desmosome. The desmosomes interlock with each other and strengthen the bond between the cells. It is interesting to note that the “spiny” nature of this layer is an artifact of the staining process. Unstained epidermis samples do not exhibit this characteristic appearance. The stratum spinosum is composed of eight to ten layers of keratinocytes, formed as a result of cell division in the stratum basale (Figure 3.13). The keratinocytes in the stratum spinosum begin the synthesis of keratin and release a water-repelling glycolipid that helps prevent water loss from the body, making the skin relatively waterproof. As new keratinocytes are produced atop the stratum basale, the keratinocytes of the stratum spinosum are pushed into the stratum granulosum. Wandering among the keratinocytes of this layer is a type of dendritic cell called the Langerhans cell, also known as a dendritic cell, which differentiates from and functions similarly to a macrophage by engulfing bacteria, foreign particles, and damaged cells that occur in this layer.
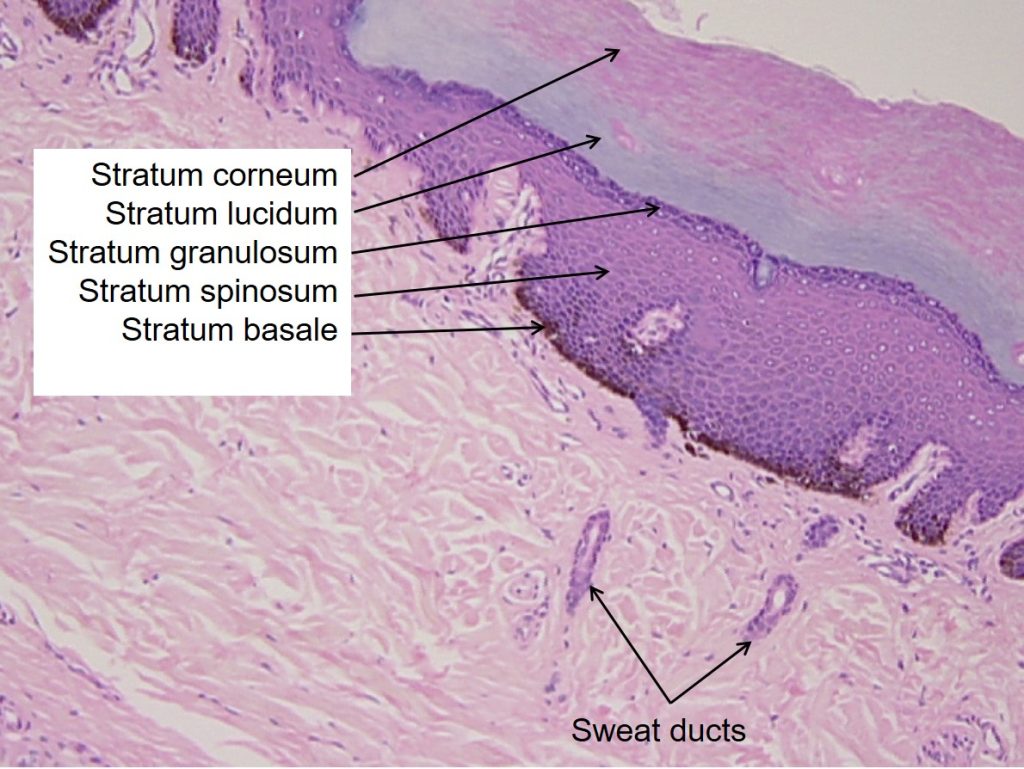
Stratum Granulosum
The stratum granulosum has a grainy appearance due to further changes to the keratinocytes as they are pushed from the stratum spinosum. The cells here (three to five layers thick) become flatter, their cell membranes thicken with a coating of lipids secreted by accumulated lamellar granules, and they generate large amounts of the fibrous protein keratin. The accumulated granules give the layer its grainy appearance. The nuclei and other cell organelles disintegrate as the cells die, leaving behind the keratin and coated cell membranes that will form the stratum lucidum, the stratum corneum, and the accessory structures of hair and nails.
Stratum Lucidum
The stratum lucidum is a smooth, seemingly translucent layer of the epidermis located just above the stratum granulosum and below the stratum corneum. This thin layer of cells is found only in the thick skin of the palms, soles, and digits. The keratinocytes that compose the stratum lucidum are dead and flattened. These cells are densely packed with a clear protein derived from keratohyalin, which gives these cells their transparent (i.e., lucid) appearance and provides a barrier to water.
Stratum Corneum
The stratum corneum is the most superficial layer of the epidermis and is the layer exposed to the outside environment (see Figure 3.12). The increased keratinization (also called cornification) of the cells in this layer gives it its name. There are usually 15 to 30 layers of cells in the stratum corneum . This dry, dead layer helps prevent the penetration of microbes and the dehydration of underlying tissues, and provides a mechanical protection against abrasion for the more delicate, underlying layers. Cells in this layer are shed periodically and are replaced by cells pushed up from the
Dermis
The dermis might be considered the “core” of the integumentary system (derma- = “skin”), as distinct from the epidermis (epi- = “upon” or “over”) and hypodermis (hypo- = “below”). It contains blood and lymph vessels, nerves, and other structures, such as hair follicles and sweat glands. The dermis is made of two layers of connective tissue, the superficial papillary layer and deep reticular layer, that compose an interconnected mesh of elastic and collagen fibers, produced by fibroblasts (Figure 3.14).
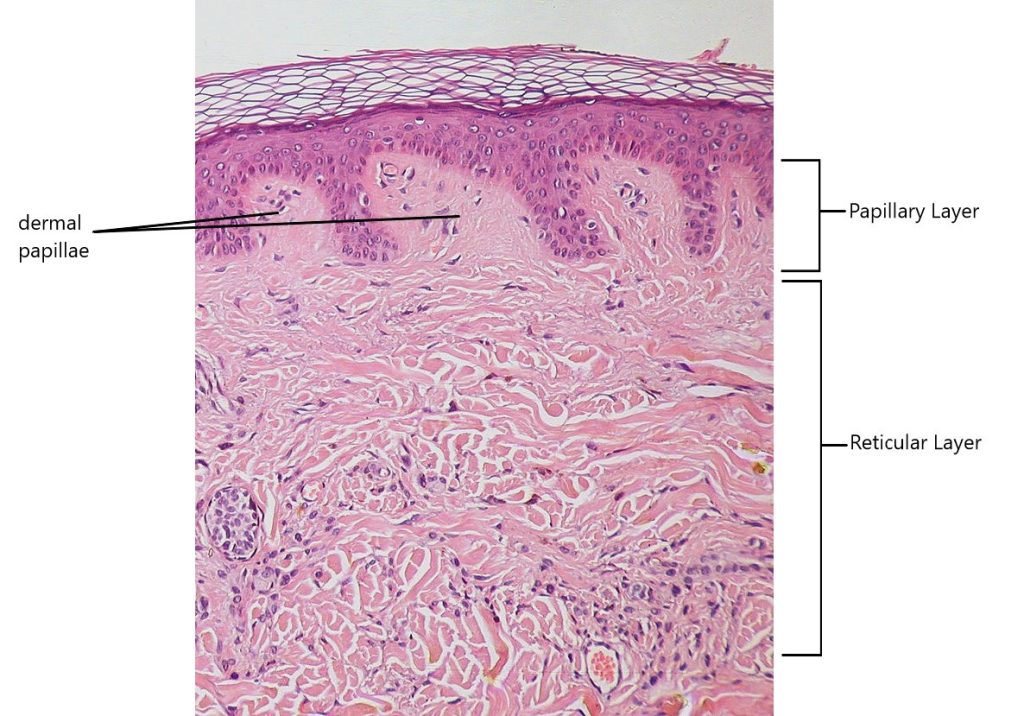
Papillary Layer
The papillary layer is made of loose, areolar connective tissue , which means the collagen and elastic fibers of this layer form a loose mesh. This superficial layer of the dermis projects into the stratum basale of the epidermis to form finger-like dermal papillae (Figure 3.14). Within the papillary layer are fibroblasts, a small number of fat cells (adipocytes), and an abundance of small blood vessels. In addition, the papillary layer contains phagocytes, defensive cells that help fight bacteria or other infections that have breached epidermis. This layer also contains lymphatic capillaries, nerve fibers, and touch receptors called tactile corpuscles (Meissner corpuscles). Tactile corpuscles can be found located in the dermal papillae, and are responsible for sensing light touch.
Reticular Layer
Underlying the papillary layer is the much thicker reticular layer, composed of dense irregular connective tissue. This layer is well vascularized and has a rich sensory and sympathetic nerve supply. The reticular layer appears reticulated (net-like) due to a tight meshwork of fibers. Elastic fibers provide some elasticity to the skin, enabling movement. Collagen fibers provide structure and tensile strength, with strands of collagen extending into both the papillary layer and the hypodermis. In addition, collagen binds water to keep the skin hydrated. Collagen injections and Retin-A creams help restore skin turgor by either introducing collagen externally or stimulating blood flow and repair of the dermis , respectively.
Hypodermis
The hypodermis (also called the subcutaneous layer or superficial fascia) is a layer directly below the dermis and serves to connect the skin to the underlying fascia (fibrous tissue) of the bones and muscles. It is not strictly a part of the skin, although the border between the hypodermis and dermis can be difficult to distinguish. The hypodermis consists of well-vascularized, loose, areolar connective tissue and adipose tissue, which functions as a mode of fat storage and provides insulation and cushioning for the skin.
Accessory and Sensory Structures of the Skin
Hair
Hair is a keratinous filament growing out of the epidermis. It is primarily made of dead, keratinized cells. Strands of hair originate in an epidermal penetration of the dermis called the hair follicle. The hair shaft is the part of the hair not anchored to the follicle, and much of this is exposed at the skin’s surface. The rest of the hair, which is anchored in the follicle, lies below the surface of the skin and is referred to as the hair root. The hair root ends deep in the dermis at the hair bulb, and includes a layer of mitotically active basal cells called the hair matrix. The hair bulb surrounds the hair papilla, which is made of connective tissue and contains blood capillaries and nerve endings from the dermis (Figure 3.15).
Just as the basal layer of the epidermis forms the layers of epidermis that get pushed to the surface as the dead skin on the surface sheds, the basal cells of the hair bulb divide and push cells outward in the hair root and shaft as the hair grows (Figure 3.16). As new cells are deposited at the hair bulb, the hair shaft is pushed through the follicle toward the surface. Keratinization is completed as the cells are pushed to the skin surface to form the shaft of hair that is externally visible. The external hair is completely dead and composed entirely of keratin. For this reason, our hair does not have sensation. Furthermore, you can cut your hair or shave without damaging the hair structure because the cut is superficial.
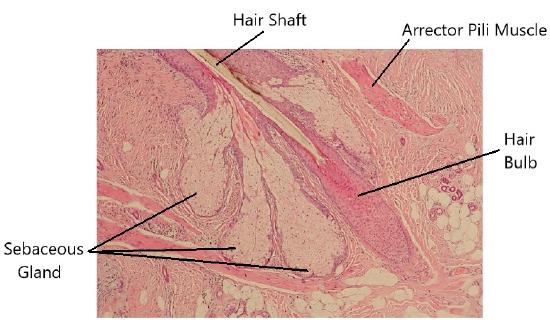
Hair serves a variety of functions, including protection, sensory input, thermoregulation, and communication. For example, hair on the head protects the skull from the sun. The hair in the nose and ears, and around the eyes (eyelashes) defends the body by trapping and excluding dust particles that may contain allergens and microbes. Hair of the eyebrows prevents sweat and other particles from dripping into and bothering the eyes. Hair also has a sensory function due to sensory innervation by a hair root plexus surrounding the base of each hair follicle. Hair is extremely sensitive to air movement or other disturbances in the environment, much more so than the skin surface. This feature is also useful for the detection of the presence of insects or other potentially damaging substances on the skin surface. Each hair root is connected to a smooth muscle called the arrector pili that contracts in response to nerve signals from the sympathetic nervous system, making the external hair shaft “stand up.” The primary purpose for this is to trap a layer of air to add insulation. This is visible in humans as goose bumps and even more obvious in animals with a heavier coat than most humans, such as dogs and cats that raise their fur when alarmed.
Sebaceous Glands
A sebaceous gland is a type of oil gland that is found all over the body and helps to lubricate and waterproof the skin and hair. Most sebaceous glands are associated with hair follicles (Figure 3.16). They generate and excrete sebum, a mixture of lipids, onto the skin surface, thereby naturally lubricating the dry and dead layer of keratinized cells of the stratum corneum, keeping it pliable. The fatty acids of sebum also have antibacterial properties, and prevent water loss from the skin in low-humidity environments. The secretion of sebum is stimulated by hormones, many of which do not become active until puberty. Thus, sebaceous glands are relatively inactive during childhood.
Sweat Glands
When the body becomes warm, sweat glands produce sweat to cool the body. Sweat glands develop from epidermal projections into the dermis. There are two types of sweat glands, each secreting slightly different products. An eccrine sweat gland is type of gland that produces a hypotonic sweat for thermoregulation. These glands are found all over the skin’s surface, but are especially abundant on the palms of the hand, the soles of the feet, and the forehead (Figure 3.17). They are coiled glands lying deep in the dermis, with the duct rising up to a pore on the skin surface, where the sweat is released. This type of sweat, released by exocytosis, is hypotonic and composed mostly of water, with some salt, antibodies, traces of metabolic waste, and dermicidin, an antimicrobial peptide. Eccrine glands are a primary component of thermoregulation in humans and thus help to maintain homeostasis.
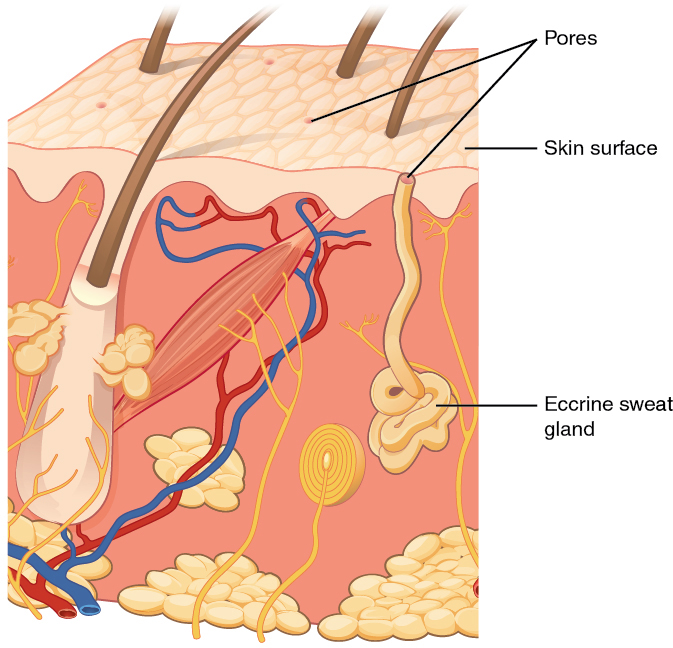
An apocrine sweat gland is usually associated with hair follicles in densely hairy areas, such as armpits and genital regions. Apocrine sweat glands are larger than eccrine sweat glands and lie deeper in the dermis, sometimes even reaching the hypodermis, with the duct normally emptying into the hair follicle. In addition to water and salts, apocrine sweat includes organic compounds that make the sweat thicker and subject to bacterial decomposition and subsequent smell. The release of this sweat is under both nervous and hormonal control, and plays a role in the poorly understood human pheromone response. Most commercial antiperspirants use an aluminum-based compound as their primary active ingredient to stop sweat. When the antiperspirant enters the sweat gland duct, the aluminum-based compounds precipitate due to a change in pH and form a physical block in the duct, which prevents sweat from coming out of the pore.
Sensory Structures
The skin acts as a sense organ because the epidermis, dermis, and the hypodermis contain specialized sensory nerve structures that detect touch, surface temperature, and pain. These receptors are more concentrated on the tips of the fingers, which are most sensitive to touch, especially the tactile (Meissner) corpuscle (Figure 3.18), which responds to light touch, and the lamellated (Pacinian) corpuscle, which responds to vibration. Tactile (Merkel) cells, seen scattered in the stratum basale, are also touch receptors. In addition to these specialized receptors, there are sensory nerves connected to each hair follicle, pain and temperature receptors scattered throughout the skin, and motor nerves innervating the arrector pili muscles and glands. This rich innervation helps you sense your environment and react accordingly.
This chapter was adapted and remixed from Douglas College Human Anatomy & Physiology I by the Douglas College Biology Department, Anatomy, Physiology, and Medical Language by NSCC, Kimberlee Carter, Marie Rutherford, and Douglas College Biology Department, Human Anatomy (OERI) by Whitney Menefee, Julie Jenks, Chiara Mazzasette, & Kim-Leiloni Nguyen, and Anatomy and Physiology (on OpenStax), by Betts, et al. and is used under a a CC BY 4.0 international license. Download and access OpenStax Anatomy and Physiology for free at https://openstax.org/books/anatomy-and-physiology/pages/1-introduction.
