
Chapter 2: Flower Morphology and Distribution
Deborah Muenchrath; Arden Campbell; Laura Merrick; Thomas Lübberstedt; Shui-Zhang Fei; and Walter Suza
Sexual Reproduction
Kinds of Flowers



Inflorescence type influences the techniques that are used to control pollination in developing cultivars and in maintaining the genetic purity of cultivars. Inflorescence types can also be used to identify plants.
Flowers are classified into a couple of categories. Flowers are either complete or incomplete and either perfect or imperfect. A flower having all of the main floral parts (sepals, petals, pistils, and stamens) is said to be complete, whereas a flower lacking one or more of these structures is said to be incomplete. The stamen (male part) and pistil (female part) are not always present together in a single flower. When both are present, the flower is said to be perfect (or bisexual). Imperfect flowers are those that are unisexual, either male or female.
| Complete flowers | Incomplete flowers |
|---|---|
| Soybean
Alfalfa Clovers Common bean Vetches Cotton  Tomato Rapeseed Sunflower Tomato Cabbage Tobacco |
Maize
Sorghum Oat Barley Wheat Sugar beet  Fig Date palm Forage grasses Turf grasses Rice Spinach |
Notice that plants in the legume family (Leguminosae or Fabaceae) have complete flowers, whereas plants belonging to the grass family (Gramineae or Poaceae) have incomplete flowers.
Flower Dissection
Dissect a complete and incomplete flower. Think about how the presence or absence of a floral structure might influence the pollination process, and thus, the methods that can be used to develop improved cultivars or to maintain the genetic purity of the cultivar.
Complete Flower

Incomplete Flower

Grass Floret

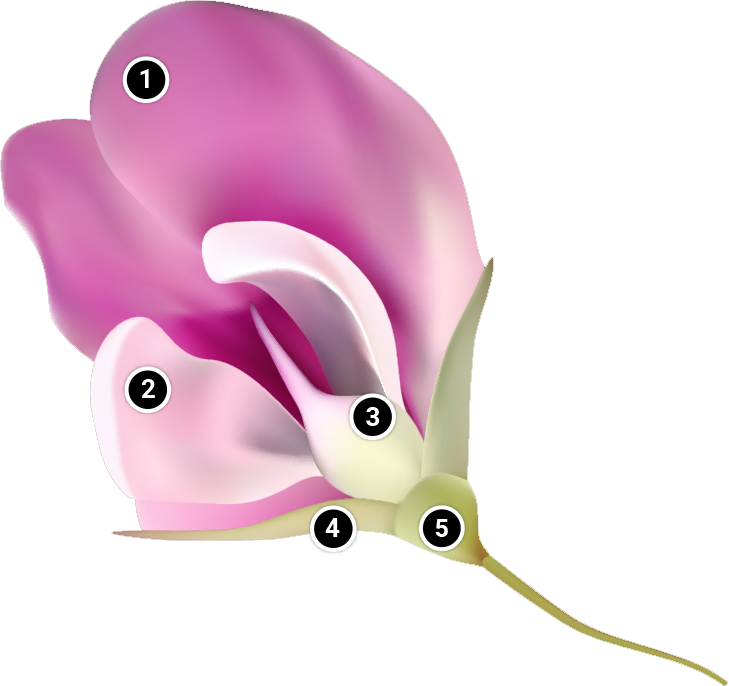
- Standard petals: Collectively, petals are called the corolla. Petals are typically large and conpicuous and are not required for reproduction. Soybean has five petals: one standard petal, two wing petals and two keel petals
- Wing petals: The dissected view of the two wing petals.
- Keel: The keel is composed of two united petals. The keel encloses the stamina column. Stamens are the pollen-bearing organs of the flower. Stamens are composed of slender stalks (filaments) that support anthers.
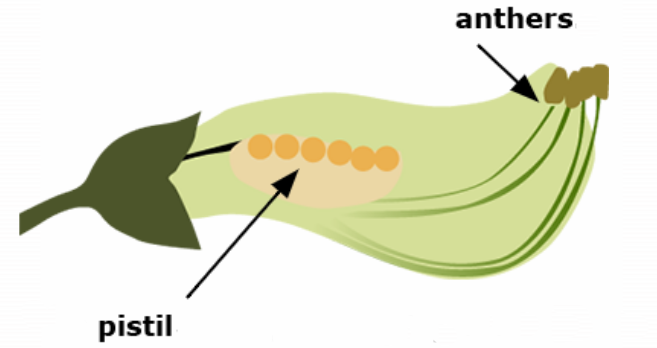 Pollen grains are produced in the anthers. The pistil is the seed-bearing organ of the flower. It consists of stigma, style, and ovaries. The stigma is the part that is receptive to pollen. Following pollination and fertilization, seed form in ovaries.
Pollen grains are produced in the anthers. The pistil is the seed-bearing organ of the flower. It consists of stigma, style, and ovaries. The stigma is the part that is receptive to pollen. Following pollination and fertilization, seed form in ovaries.
- Sepals: Like the petals, sepals are not neccessary for reproduction. Sepals are small and inconspicuous. They enclose and protect the flower while still a bud. Collectively they form the calyx.
- Pedicel: The pedicel is the stalk of the flower, attaching to the plant.
Select the floral part or parts necessary for reproduction:
| Flower Characteristics | Terms | Examples |
|---|---|---|
| Male and female expression in INDIVIDUAL FLOWERS | ||
| Male and female in ONE flower | bisexual, hermaphroditic, monoclinous, perfect | Wheat, peach |
|
protandry (prevent self-pollination) | Carrot, walnut |
|
protogyny (prevent self-pollination) | Pearl millet, pecan |
|
chasmogamy (promote self-pollination) | Violet, rye |
|
cleistogamy (ensure self-pollination) | Oat, peanut |
| Perfect flowers of TWO types on SAME plant | heterostyly | Buckwheat, flax |
|
pin flower | |
|
thrum flower | |
| Male and female in SEPARATE flowers | unisexual, diclinous, imperfect | Cucumber, hemp |
|
male, staminate | |
|
female, pistillate, carpellate | |
| Flower DISTRIBUTION on PLANTS | ||
| Male and female flowers on one plant | monoecious | Maize, oak |
| Male and female flowers on separate plants | dioecious | Yams, asparagus |
|
mixed, polygamous | Red maple, papaya |
|
polygamomonoecious | Coconut, mango |
|
polygamodioecious | Strawberry, holly |
Perfect and Imperfect Flowers
Perfect flowers have both staminate and pistillate structures in the same flower.
Imperfect flowers are either staminate or pistillate. An imperfect flower is staminate if it possesses stamen. Conversely, an imperfect flower is pistillate if it bears a pistil. Staminate flowers are considered “male” because they produce pollen, whereas pistillate flowers are “female” because they possess ovules. Staminate and pistillate flowers may occur on the same or different plants of the same species.
Species having such specializations are either:
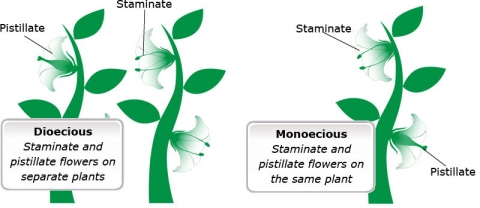
- monoecious — staminate and pistillate flowers are separate but occur on the same plant; or
- dioecious — staminate and pistillate flowers are on separate plants.
Analogous to the separate sexes in animals, a dioecious plant must have a partner of the opposite type to complete its life cycle. Usually, about half of all individuals of a dioecious species are of each type, staminate or pistillate. Thus, the dioecious condition is reproductively expensive in that only about half of the species’ plants can produce seed.
| Monoecious | Dioecious |
|---|---|
Maize
 Walnut Oil palm Squash Cassava Wile rice Castor bean White pine |
Hemp
 Hops Spinach Yam Date palm Cottonwood Asparagus Nutmeg |
The “mono-” prefix indicates one and the “di-” prefix indicates two. The “-oecious” part of the word translates to “house.” Thus, an easy way to remember the distinction between these terms is to remember that in monoecious species, the staminate and pistillate flowers reside in the same house or plant, whereas in dioecious species, these flowers reside in two different houses or plants.
Pollination and Fertilization
Pollination occurs when a pollen grain (from the staminate flower) is placed on a receptive stigma (of the pistillate flower), either naturally or artificially. Fertilization requires that a male gamete and a female gamete fuse to form a zygote. These gametes may be from the same or different plants.
There are two kinds of pollination processes in sexual reproduction.
- Self-pollination — seeds develop from the union of male and female gametes produced on the same plant or clone. The development of seed by self-pollination is also referred to as autogamy.
- Cross-pollination — seeds develop from the fusion of gametes produced on different plants. The development of seed by cross-pollination is known as allogamy.
Self-Pollination
Several floral mechanisms enforce self-pollination.
- Flowers do not open, preventing external pollen from reaching the stigma.
- Anthesis occurs before the flower opens.
- Stigma elongates through the staminal column (filaments and anthers) immediately after anthesis.
- Floral organs may obscure the stigma after the flower opens.
Although these mechanisms usually enforce self-pollination, a low frequency of cross-pollination may occur. The frequency of cross-pollination in normally self-pollinating species generally depends on the species and environmental conditions.
Soybean is an example of a species that is normally self-pollinated. Before the flower opens, the anthers burst and pollen grains fall out of the anthers on to the receptive stigma contained in the same flower: self-pollination occurs.
Cross-Pollination
Floral Mechanisms of Promotion
Several floral mechanisms promote cross-pollination.
- Emergence or maturity of the staminate and pistillate flowers is asynchronous.
- Flowers are monoecious or dioecious. Mechanical obstruction between the staminate and pistillate flowers in the same individual prevents self-pollination. Gametes produced on the same plant or clone are unable to effect fertilization.

- Mechanical obstruction between the staminate and pistillate flowers in the same individual prevents self-pollination.

Alfalfa flowers, for example, have a membrane over the stigma that precludes self-pollination. When a bee lands on the flower, the keel is tripped, rupturing the membrane and exposing the stigma to pollen carried by the bee from other plants it has visited, effecting cross-pollination.
- Gametes produced on the same plant or clone are unable to effect fertilization.
- Self-sterility — gametes from same individual cannot successfully fuse to form a zygote. Sterility can be caused by lack of function of pollen (male gametes) or ovules (female gametes).
- Male sterility — either genetic or cytoplasmic, occurs because the pollen is not viable. Female sterility occurs when the ovule is defective or seed development is inhibited.
- Self-incompatibility — self-pollination may occur, but fertilization and seed set fail.
Pollen Transportation
Pollen is transported from the staminate flower to the pistillate flower by wind, insects, or animals. Occasionally pollen is transported to receptive stigma of the same individual and self-pollination may occur. For example, pollen from the tassel of a maize plant may land on and pollinate silks on the same plant, effecting self-pollination.
Sunflower is ordinarily cross-pollinated. Bees often carry pollen from one plant and deposit it on other plants.


Classification
Plants are classified as either self- or cross-pollinated based on which of these processes most frequently produces its seed. Click each category for more information.
You encounter an unfamiliar flowering plant. What key floral feature(s) would you check to determine the plant’s likely mode of pollination, self or cross-pollinating?
For each of the following types, indicate the probable mode of pollination by clicking on the appropriate button. Assume no male sterility or self-incompatibility.
Asexual Reproduction
Some species can be propagated without a gametophytic stage. The fusion of gametes (fertilization) is omitted from the life cycle. Reduction in chromosome number (meiosis) and seed production may or may not occur. Asexual reproduction produces individuals genetically identical to the maternal parent.
There are several mechanisms of asexual reproduction.
- Vegetative Propagation
- Tissue Culture
- Apomixis
Vegetative Propagation
In some species, new individuals can arise from a group of differentiated or undifferentiated cells of the parent plant; no embryo or seed is produced. Because such new individuals develop asexually from a single parent, they are genetically identical to that parent. These progeny are clones. Numerous tissues and organs may asexually produce progeny.
- Rhizomes – Rhizomes are specialized underground stems that can branch at nodes to produce new plants. Banana, bromegrass, hops, and johnsongrass can be reproduced from rhizomes.
- Stolons – These “runners” or horizontal-growing, above-ground stems develop adventitious roots whose axillary buds can become independent plants. Strawberry is an example of a crop that can be reproduced from stolons.
- Bulbs and bulbils – These short underground stems have thickened or fleshy scales (modified leaves) that can form buds. These buds detach and form “offsets” or new individuals. Onions and garlic are commonly propagated from bulbs.
- Tubers – Tubers are also short, enlarged stem tissue, containing food reserves. Nodes or “eyes” in such tissue can give rise to adventitious roots and separate plants. Potatoes are commonly propagated from eyes cut from tubers.
- Suckers – Suckers arising as lateral shoots from the base of stems can separate and form new plants. Pineapple, sweet potato, and date palm are examples. Suckers may also derive from adventitious buds on the roots. Roses, poplars, and some other woody species can be propagated from such root cuttings or rootstocks.
- Corms – A corm is an underground, tuber-like base of a vertical stem that can also produce a separate plant. Taro, an important starch crop in Southeast Asia and the Pacific Islands, is propagated from corms. Banana also can be propagated from corms.
- Stem cuttings – When placed in moist soil, cuttings from aerial stems of some species, such as sugarcane, pineapple, and cassava, can give rise to new plants from the nodes and lateral buds.
The usual mode of reproduction of some species is vegetative. However, other species that reproduce sexually are more commonly propagated vegetatively to maintain genetic purity, including some forage cultivars and many horticultural species.
Vegetative reproduction does not usually provide opportunity for selection of genetic variants.
Tissue Culture
Tissue culture is a specialized type of asexual propagation. Tissue culture usually involves excision of undifferentiated cells or meristematic pieces of a plant and growing these in vitro on sterile nutrient agar medium; cell division is by mitosis. By manipulating the components of the medium, the tissue can be prompted to develop roots or shoots. Eventually, new individuals may be separated and transplanted to soil.
Tissue culturing takes advantage of the totipotency of somatic cells. That is, these cells contain the plant’s entire genome and have the potential to develop into whole plants. Some species that cannot normally be reproduced vegetatively may be reproduced by tissue culture.
Tissue culture is of interest to plant breeders as a technique to
- maintain and propagate genetically identical plants that otherwise can only be reproduced sexually;
- provide disease-free plants of species that often transmit pathogens to progeny when propagated by conventional vegetative means; and
- create novel genetic variation within which selections can be made. Under some conditions, tissue culturing can promote genetic changes.
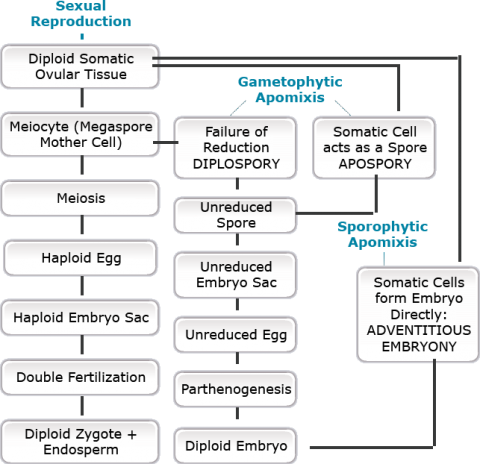
Apomixis Process
Apomixis differs from other forms of asexual reproduction in that seed is produced. Unlike sexual reproduction, however, apomictic seed is developed from sexual organs or related structures without fertilization. Pollination is also usually omitted.
Agamospermy
Apomixis generally involves forms of agamospermy, which is a process through which seeds develop without fertilization. There are two different degrees of agamospermy.
- Obligate — Seed produced arises from asexual reproduction.
- Advantages: Preserves genotype, including heterozygotic genotypes
- Disadvantages: Precludes genetic recombination and variation for selection of improved cultivars
- Facultative — Although most of the seed generated is asexually produced, sexual reproduction occurs regularly.
- Advantages: Permits development of genetic variation for selection of improved cultivars
- Disadvantages: Cultivars may be genetically unstable, making it difficult to maintain the desired genotype
Each of these degrees of agamospermy provides advantages and disadvantages from the plant breeding perspective.
There are also two general types of agamospermy.
- Autonomous — Endosperm forms without pollination or fertilization.
- Pseudogamous — Although fertilization (the fusion of gametes) does not occur, pollination is apparently required to stimulate apomictic embryo or embryo sac development to produce seed. Pollination adds no genetic material.
Mechanisms of Cause
The mechanisms that cause apomixis differ by the cell that undergoes mitosis to produce the embryo of the seed.
- Adventitious embryony — The embryo develops directly from diploid sporophytic tissue, skipping the gametophytic stage. This is the simplest form of agamospermy.
- Apospory — Nucellus or integument cells, which are somatic cells, undergo mitosis to produce a diploid embryo sac.
- Apospory is the most common form of apomixis in angiosperms.
- Diplospory — The embryo and endosperm derive from the diploid megaspore mother cell. The megaspore mother cell’s nucleus divides by mitosis, rather than meiosis, resulting in a diploid embryo sac.
- Parthenogenesis — The egg cell divides mitotically to form the embryo without fertilization.
- Androgenesis — A haploid embryo develops from a male sperm nucleus after it enters the embryo sac. The individual that develops from the seed is haploid and has the genotype of the sperm from which it is derived.
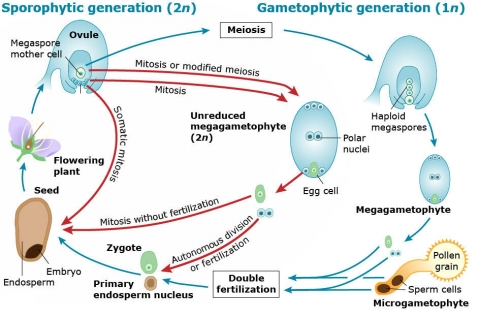
Apomictic embryos may form from reduced (haploid) or unreduced (diploid) cells. For each situation, select the button according to whether the resulting embryo could be homozygous, heterozygous, neither, or both. Answer both parts of this question and then check your answer.
Discussion
Crops can be self-pollinated, cross-pollinated, or vegetatively propagated. Discuss the breeding consequences of these three different methods of propagation. In addition: previously, a student suggested that with today’s technologies, plants can simply be converted into self- or cross-pollinated or into vegetatively propagated species. Do you agree? Provide arguments in favor or against this statement, and examples, in case you are aware of any. Finally, if it was possible, which type of crops would be your favorite, and why?
For each of the following terms, identify whether the term is associated with sexual, asexual, or both manners of reproduction by clicking on the appropriate button.
References
Bicknell, A. R. and Koltunow, M., A. 2004. Understanding Apomixis: Recent Advances and Remaining Conundrums [Online]. Available by web link to American Society of Plant Biologists (http://www.plantcell.org/content/16/suppl_1/S228.full.pdf) (verified Jan 24, 2017).
Bashaw, E.C. 1980. American Society of Agronomy-Crop Science Society of America: 1980 Madison, WI, Hybridization of Crop Plants [Online]. Available at http://www.agron.iastate.edu/faculty/fehr/HOCP/03HOCP.pdf (verified Jan 24, 2017).
Chang, M.T., and M.G. Neuffer. 1989. Maize microsporogenesis. Genome 32(2): 232-244.
Lersten, N. R. 1980. Reproduction and Seed Development. Chapter 2, p. 17-44. In W.R. Fehr. and H. H. Hadley, editors. Hybridization of Crop Plants. American Society of Agronomy, Madison, WI.
Ozias-Akins, P. 2006. Apomixis: Developmental Characteristics and Genetics. Critical Reviews of Plant Sciences. 25: 199-214.
Pierce, B. A. 2008. Genetics: A Conceptual Approach. 3rd edition. W.H. Freeman, New York.
Schlegel, R.H.J. 2010. Dictionary of Plant Breeding. 2nd edition. CRC Press, New York.
Simmonds, N. W., and J. Smartt. 1999. Principles of Crop Improvement. 2nd. edition. Blackwell Science, Oxford, UK.
Smartt, J., and N. W. Simmonds. 1995. Evolution of Crop Plants. 2nd. edition. Longman, Essex, UK.
Vielle-Calzada, J-P., C. F. Crane and D. M. Stelly. 1996. Apomixis: the asexual revolution. Science 274: 1322-1323.
mcar2010. 2010. Plant reproduction.mov [Online]. YouTube. Available at http://www.youtube.com/watch?v=ZvBMQIrlhYw (verified Jan 24, 2017).
That variability in an experiment that is random or cannot be explained.
A term that refers to the metabolism of a living organism and the array of biochemical pathways and processes
(1) Plants comprising the population are genetically identical.
(2) Population is comprised of genetically identical plants.
Condition that exists in a population of individuals with the same genotype.
A term that refers to the metabolism of a living organism and the array of biochemical pathways and processes
A transformation useful on data collected of proportions or percentages. These data may be transformed by taking the inverse of the sine or arcsine of the number.
A localized group of actively dividing cells from which permanent specialized tissues differentiate.
The donor plants in the case of DH are the F1 or F2 progeny resulting from a breeding cross from which homozygous lines are desired.
Polyploid in which all chromosomes of one homologous type carry the same recessive allele for the particular gene.
